Title: Traditional Chinese medicine describes the pathways of signaling molecules
Authors: Yang LIU*
Affiliations:
The Hong Kong Polytechnic University, Hung Hom, Kowloon, Hong Kong.
- To whom correspondence should be addressed: mmyliu@polyu.edu.hk
Abstract
The pathways of signaling molecules are important to understanding how signaling molecules regulate physiological function and also in predicting the pathological development which is important to therapeutic strategy, however the thorough knowledge of these pathways is still lack. Traditional Chinese medicine (TCM) used Yang and Yin to describe the physiological processes in human body, but its scientific basis is unknown. Here we establish the direct mapping between Yang-Yin and signaling molecules. According to “Yellow Emperor’s Inner Canon” (YEIC), the fundamental doctrinal source for TCM, there are five groups of Yang and Yin, each has particular physiological functions and the different groups have particular relationships. We used the iterative selection method to compare these physiological functions with the functions of signaling molecules, and establish the direct mapping between Yang-Yin and signaling molecules. With the mapping, it is shown that the pathways of these signaling molecules are the same as the dialectical relationships of these Yang-Yins and agree well with TCM therapeutic strategies. The Chinese herbal medicine actually regulates the level of corresponding signaling molecule. With the mapping, it is clearly shown that the modern medicine and TCM identify the same pathogenesis of diseases, such as hematuria, constipation, hemorrhoidal disease, nocturnal enuresis, frequent urination, gall stone, acute cholecystitis and hypertension; and adopt the same treatment strategy. Modern medicine and TCM describes the same physiological process using different terminology.
Key words: Signaling molecules, traditional Chinese medicine, pathways, Yang and Yin, therapeutic strategy.
Glossary
| AKR | aldo-keto reductase, Liver Yin in TCM |
| Ang | Angiotensin, Heart Yin in TCM |
| ANP | atrial natriuretic peptide, Spleen Yang in TCM |
| EPO | Erythropoietin, Liver Yang in TCM |
| FGF7 | fibroblast growth factor-7, Lung Yang in TCM |
| Five Phases | TCM term, includes heart phase, spleen phase, lung phase, kidney phase, liver phase |
| GSH | Glutathione, Liver Qi in TCM |
| Heart phase | TCM term, includes heart organ, Heart Yang, Heart Yin, and Heart Qi |
| HGF | hepatocyte growth factor, Liver Yang in TCM |
| HIF | hypoxia inducible factor, Lung Qi in TCM |
| IGF | insulin-like growth factor, Heart Yang in TCM |
| Kidney phase | TCM term, includes kidney organ, Kidney Yang, Kidney Yin, and Kidney Qi |
| Liver phase | TCM term, includes liver organ, Liver Yang, Liver Yin, and Liver Qi |
| Lung phase | TCM term, includes lung organ, Lung Yang, Lung Yin, and Lung Qi |
| Mg | Magnesium, Heart Qi in TCM |
| NO | nitric oxide, Kidney Qi in TCM |
| PTHrP | parathyroid hormone-related protein, Kidney Yin in TCM |
| Qi | substance in TCM, has function of activation and regulation |
| SOD | superoxide dismutase, Liver Yin in TCM |
| Spleen phase | TCM term, includes spleen organ, Spleen Yang, Spleen Yin, and Spleen Qi |
| TCM | traditional Chinese medicine |
| VEGF | vascular endothelial growth factor, Lung Yang in TCM |
| Yang | Key substance in TCM, has function of warming/developing/proliferation |
| YE-81 | Yellow Emperor’s Classic on 81 Medical Problems |
| YEIC | Yellow Emperor’s Inner Canon |
| Yin | Key substance in TCM, has function of cooling and astringent |
Introduction
The pathways of signaling molecules are important to understanding how signaling molecules regulate physiological function and also in predicting the pathological development which is important to therapeutic strategy. However, despite the documented importance of signaling molecules to human life, we still lack thorough knowledge of pathways of signaling molecules.
Traditional Chinese medicine (TCM) has been effective for thousands of years in China. Ancient TCM physicians believed that there existed key substances to regulate the physiological processes and named them as Yang and Yin, where Yang has warming/developing function and Yin has cooling/astringent function. Yang and Yin are contrary but interdependent in human body. According to the physiological functions of these key substances, ancient TCM physicians categorized these Yang and Yin into heart phase, spleen phase, lung phase, kidney phase and liver phase, i.e., the Five Phases. Besides Yang and Yin, TCM physicians also found there existed another substance in each organ whose function is actuation/regulation, and they named it as Qi. Thus, each organ phase includes the organ itself, and the organ’s Yang, Yin and Qi. According to “Yellow Emperor’s Inner Canon” (YEIC) [1], the fundamental doctrinal source for TCM, each organ phase has particular physiological functions, and the Five Phases regulate each other and follow the “mother-son” and “master-servant” relationships (Figure 1). Therefore, TCM physicians developed diagnosis theories and therapeutic strategies based on these dialectical interrelationships.
However, there is an obstacle to developing a general frame work linking TCM theory and modern physiology, i.e., one cannot scientifically identify Yang, Yin and Qi of each organ. Due to the tremendous studies in recent years on signaling molecules, we have an unprecedented opportunity to explore the link between TCM theory and life science. Here we use the iterative selection method to establish the direct mapping between these Yang-Yin and signaling molecules, and have three central findings. First, the Yang or Yin corresponds to a type of signaling molecules. Second, the pathways of these signaling molecules are the same as the dialectical relationships of these Yang and Yin and agree well with TCM therapeutic strategies. Third, the modern medicine and TCM identify the same pathogenesis of diseases, such as hematuria, constipation, hemorrhoidal disease, nocturnal enuresis, frequent urination, gall stone, acute cholecystitis and hypertension; and adopt the same therapeutic strategy. The modern medicine and TCM describe the same physiological processes using different terminology.
Iterative Selection Method
Using the functions of Heart phase, Spleen phase, Lung phase, Kidney phase and Liver phase in TCM as constraint conditions, we apply the following Iterative Selection method to identify the signaling molecules. Suppose the signaling molecules form the following set,
X = {x: signaling molecules}, and Heart phase, Spleen phase, Lung phase, Kidney phase and Liver phase are A, B, C, D, E, respectively.
Step 1: identify the signaling molecules in each organ
There are four constraint conditions in each organ as shown in Figure 1.
φA(u1, u2, u3, u4) denotes the four conditions in Heart phase, where
u1: the molecule should have significant effect on heart,
u2: the molecule governs blood vessel,
u3: the molecule is enlightened in tongue,
u4: the molecule affects the appearance of face [2].
φB(u1, u2, u3, u4) denotes the four conditions in Spleen phase, where
u1: the molecule should have significant effect on spleen,
u2: the molecule governs muscle,
u3: the molecule is enlightened in mouth,
u4: the molecule affects the appearance of lips.
φC(u1, u2, u3, u4) denotes the four conditions in Lung phase, where
u1: the molecule should have significant effect on lung,
u2: the molecule governs hair and skin,
u3: the molecule is enlightened in nose,
u4: the molecule affects the appearance of body hairs.
φD(u1, u2, u3, u4) denotes the four conditions in Kidney phase, where
u1: the molecule should have significant effect on kidney,
u2: the molecule governs bone,
u3: the molecule is enlightened in ears,
u4: the molecule affects the appearance of hairs.
φE(u1, u2, u3, u4) denotes the four conditions in Liver phase, where
u1: the molecule should have significant effect on liver,
u2: the molecule governs tendon,
u3: the molecule is enlightened in eyes,
u4: the molecule affects the appearance of paws.
Then, compare X with φ, and obtain the class Y,
Y = {x: φj(u1, u2, u3, u4)}, for Heart phase, Spleen phase, Lung phase, Kidney phase and Liver phase, respectively.
Step 2: identify Yang, Yin and Qi for each organ
In TCM, Yang has developing/warming effect, Yin has astringent/cooling effect, and Qi has the function of activation/regulation. These constraint conditions are set as Φ1, Φ2, and Φ3. Then, compare class Y with Φ1, Φ2, and Φ3 respectively, and obtain either Yang, or Yin, or Qi for each organ phase:
A1 = y: Φ1, A2 = y: Φ2, and A3 = y: Φ3;
B1 = y: Φ1, B2 = y: Φ2, and B3 = y: Φ3;
C1 = y: Φ1, C2 = y: Φ2, and C3 = y: Φ3;
D1 = y: Φ1, D2 = y: Φ2, and D3 = y: Φ3;
E1 = y: Φ1, E2 = y: Φ2, and E3 = y: Φ3.
Step 3: refine the selection according to “mother-son relationship” in Figure 1
Since Yang has developing effect, according to the “mother-son” relationship, the organ Yang should be secreted from the “mother” organ, e.g., Kidney Yang should be secreted from lung. We use this condition to refine the A, B, C, D, E in Step 2.
Step 4: refine the selection by comparing pathways
We compare the pathways of signaling molecules with the interrelationships of the Five Phases in Figure 1 to further refine the selection in Step 3.
Step 5: validate the selection based on Chinese herbal medicine
There are some clinical and experimental data to show that some TCM herbal medicine would regulate the signaling molecules, and we use these available data to validate the results in Step 4.
Results
The mapping between Yang-Yin-Qi and Signaling molecules
According to the “Yellow Emperor’s Inner Canon” (YEIC) [1], the Heart phase should be generated in liver organ, govern blood vessel, and be enlightened in tongue; and its prosperity is reflected by face [2]. The Spleen phase should be generated in heart organ, govern muscle, and be enlightened in mouth; and its prosperity is reflected by lips. The Lung phase should be secreted in spleen organ, govern skin and hair, and be enlightened in nose; and its prosperity is reflected in body hair. The Kidney phase should be generated in lung organ, govern bone, be enlightened in ears, and affect the appearance of hair. The Liver phase should be generated in kidney organ, govern tendon, be enlightened in eyes, and affect the appearance of “paw” (finger, toe, nail). Using these physiological functions as constraint conditions, we apply the iterative selection method to establish the direct mapping between the signaling molecules and the Yang-Yin-Qi in Five Phases as tabulated in Table 1 and discussed in Supplementary Information.
The consistency between pathways of signaling molecules and dialectical relationships of Five Phases in TCM
Figure 2 shows that the signaling molecules corresponding to Yang-Yin-Qin within the same phase regulate each other (Supplementary Information), and the pathways of signaling molecules among different phases are consistent with the dialectical relationships of Five Phases in TCM. Some of these dialectical relationships have been widely applied in TCM therapies for hundreds of years, and Figure 3 demonstrates that these typical TCM therapeutic strategies are highly consistent with the pathways of those signaling molecules.
In Figure 1, the five arrows along the diagonal direction denote the “master-servant” relationships. We show that each relationship is consistent with the pathway of signaling molecules. YEIC states that Spleen phase is subjected to Liver phase. “Yellow Emperor’s Classic on 81 Medical Problems”, another fundamental doctrinal source for Chinese medicine on therapeutic strategies, states that the disease in liver transfers to spleen, to treat the disease in liver, the best physician should enhance Spleen phase, and then the disease in liver would be recovered [3]; in physiological reality, circulating retinoic acid concentrations are lower in subjects with nonalcoholic fatty liver disease and are associated with hepatic lipid metabolism [4], and ghrelin treatment increases GSH levels[5] and activity of SOD [6]. Moreover, EPO stimulates ANP secretion [7].
YEIC denotes that Kidney phase is subjected to Spleen phase. The popular guidance in TCM therapy stated that Kidney phase is dependent on Spleen phase and vice versa[8]; in physiological reality, ANP increases calcitonin gene-related peptide (CGRP) within human circulation [9] and CGRP stimulates ANP secretion [10].
YEIC denotes that Heart phase is subjected to Kidney phase. The popular guidance in TCM therapy stated that Heart phase and Kidney phase are in harmony, to enhance Heart phase, one should strengthen Kidney phase; to enhance Kidney Yin, one should lower down Heart Yang [11]. In physiological reality, calcitonin increases concentration of IGF [12], and IGF signaling antagonizes the Wnt pathway [13].
YEIC denotes that Lung phase is under the dominion of Heart phase. In physiological reality, magnesium deficiency contributes to pulmonary complications [14]; IGF-1 induces lung fibroblast activation [15]; IGF-1 induces VEGF [16], exogenous administered FGF7 cDNA causes increases in IGF-1 [17], and the combination of IGF-1 and FGF7 cDNA significantly improves dermal and epidermal regeneration [18]; and Ang II increases FGF7 mRNA levels [19] and stimulates VEGF synthesis [20].
YEIC denotes that Liver phase is subjected to Lung phase. In physiological reality, it was found that FGF7 promotes liver regeneration [21]; VEGF promotes proliferation of hepatocytes through reconstruction of liver sinusoids by proliferation of sinusoidal endothelial cells [22]; vitamin C deficiency promotes fatty liver disease development [23] and vitamin C plus E combination treatment is a safe and effective treatment option in patients with fatty liver disease [24]; and HIF plays a role across a range of hepatic pathophysiology [25]. Moreover, VEGF is crucial for EPO-induced improvement of cardiac function [26]; vitamin C elevates red blood cell glutathione in healthy adults [27]; HIF induces EPO production [28] and induction of HIF-1 varies with the GSSG/GSH ratio [29]; FGF-7 stimulates HGF mRNA and protein expression in ovarian surface epithelium [2].
The five arrows along the outer circle in Figure 1 denote the “mother-son” relationships. The signaling molecule corresponding to Yang in each phase is secreted/generated in “mother” organ (Supplementary Information). Moreover, the popular guidance in TCM therapy stated that “Liver and kidney have the same origin and they should be treated together” [30]; in physiological reality, EPO is produced principally in the liver during fetal gestation and in the kidney after birth, and in response to anemia, EPO gene transcription is markedly induced in the kidney and liver [31].
For the relationship of Lung and Spleen phases, TCM stated that “to treat lung disease, one should treat spleen, as Spleen phase can promote Lung phase” [30], and the popular TCM therapy strategy of enhancing Lung Qi is to enhance Spleen phase; in physiological reality, HIF-1α and VEGF mRNAs are significantly increased after ghrelin administration [32]; retinoic acid increases HIF-1α [33] and markedly stimulates FGF7 expression [34].
TCM considers that Lung phase and Kidney phase are mutually promoted, and the Kidney Yin can be increased by enhancing lung Qi [11]. In physiological reality, HIF modulates Wnt/β-catenin signaling [35], induces PTHrP [36], and is regulated by NO [37]; Wnt activates FGF7 during epidermal stratification [38], and regulates vitamin C biosynthesis [39]; and vitamin C increases nitric oxide synthase activity [40].
TCM denotes that Liver phase increases Heart phase, in physiological reality GSH enhances intracellular Mg both in vivo and in vitro [41]. Moreover, Heart phase enhances Spleen phase in TCM, and it was found that IGF-II increases the protein level of ANP [42] (Figures 2 & 3).
The function of TCM herbal medicines and formula is to regulate signaling molecules
TCM herbal medicines and formula have been widely applied in TCM therapies to regulate Yang-Yin-Qi in each organ phase. We validated our direct mapping by comparing the functions of TCM herbal medicines with the effects of these herbal medicines on signaling molecules (Supplementary Information).
Discussion
With the direct mapping, we can demonstrate that TCM and modern medicine actually identify the same pathogenesis of human disease and apply the same treatment strategy. TCM considers that heart is connected with small intestine. Yuan-fang Chao (550-630) was a famous imperial doctor in Chinese history, he stated that “As heart is connected with small intestine, if heart has excess heat (excess Heart Yang), then the heart heat stagnates at small intestine and results in hematuria” [43], and the effective TCM treatment strategy is to lower the Heart Yang (or IGF). In physiological reality, IGF-1 level was higher in Henoch-Schönlein purpura patients with hematuria than in controls [44], and the angiotensin-converting enzyme inhibitor can effectively treat gross hematuria [45]. As angiotensin-converting enzyme inhibitor therapy significantly reduces circulating IGF-1 [46], this treatment strategy of modern medicine is the same as that of TCM.
TCM considers that spleen is connected with stomach. Dong-yuan Li (1180-1251), a Chinese physician famous in the relationship between spleen and stomach, stated that raising Spleen Yang (or ANP) could benefit stomach [47]. In physiological reality, endogenous ANP stimulates somatostatin secretion from antrum of human stomach, and stimulation of somatostatin secretion is coupled to inhibition of gastrin secretion in the antrum [48]. Moreover, ghrelin stimulates gastric motility [49].
TCM considers that lung is connected with large intestine. Zonghai Tang (1851-1908) stated that “Lung Yang moves heat to large intestine thus make it stagnated, and insufficient Lung Yin (or vitamin C) results in constipation” [50]. Shi-zhen Li (1518-1593), a famous Chinese physician in history and the author of Compendium of Materia Medica, suggested using ficus carica to treat hemorrhoidal disease [51]. In physiological reality, higher expression of VEGF exists in hemorrhoidal tissue [52], and it was found that constipated children had significantly lower intakes of vitamin C [53]. Moreover, ficus carica decreases VEGF level significantly [54] and is an excellent source of ascorbic acid [55]. These indicate that the pathogenesis and treatment strategy of modern medicine are the same as those of TCM.
TCM considers that kidney is connected with bladder. Yuan-fang Chao (550-630) stated that kidney and bladder controls micturition, and deficient Kidney Qi (or NO) causes frequent urination and nocturnal enuresis [43]. In physiological reality, NO synthase plays a functional role for controlling micturition [56], neuronal NO synthase gene polymorphism may be associated with primary nocturnal enuresis in children [57], and Nicorandil, a nitric oxide donor, improves the frequent urination [58]. These are consistent with the statement of TCM.
TCM considers that liver is connected with gallbladder. TCM considers that one of the pathogenesis of gall stone is Liver Qi melancholy, and one of the pathogenesis of cholecystitis is Liver Qi (or GSH) stagnation [59]. In physiological reality, gall stone patients have altered gallbladder functions and their GSH levels are lower than those of gall stone free subjects [60]; in acute cholecystitis patients, the total glutathione in blood is lower while the activity of glutathione metabolism enzymes in red blood cells is inhibited [61]. The pathogenesis is consistent with that of TCM.
Taking hypertension as another example. TCM physicians consider that one of the pathogeneses of hypertension is “Liver Yang is hyperactive” [62]; in physiological reality, chronic administration of EPO induces hypertension [63], and serum HGF concentration in hypertensive patients is significantly higher than that in normal subjects [64]. TCM considers another pathogenesis of hypertension is “Both Kidney Yin and Liver Yin are deficient” [62]; in physiological reality, hypertension patients had low or undetectable levels of plasma renin activity [65], localized loss of pulmonary artery extracellular SOD augments chronic hypoxic pulmonary hypertension [66], AKR activity is significantly low in spontaneously hypertensive rat medulla [67], and hypertension is associated with downregulation of canonical Wnt signaling pathways [68]. All of these examples indicate that TCM and modern medical science are describing the same thing but using different terminology!
The direct mapping between TCM and signaling molecules enables us to investigate the effect of balance of signaling molecules on human health and understand the closed-loop of pathways of these molecules, that would lead to new therapeutic strategies and insight into the rules governing physiological processes.
Table 1 Corresponding signaling molecules of each organ phase
| Yang | Yin | Qi | |
| Heart phase | IGF | Ang | Mg |
| Spleen phase | ANP | aldosterone,
retinoic acid |
ghrelin |
| Lung phase | FGF7, VEGF | ascorbic acid | HIF |
| Kidney phase | calcitonin, PTHrP | Wnt | NO |
| Liver phase | EPO, HGF | renin, SOD, AKR | GSH |
Figure Legends
Figure 1 The functions and interrelationships of the Five Phases in TCM. The Heart phase should be generated in liver, govern blood vessel, and be enlightened in tongue; and its prosperity is reflected by face [2]. The Spleen phase should be generated in heart, govern muscle, and be enlightened in mouth; and its prosperity is reflected by lips. The Lung phase should be secreted in spleen, govern skin and hair, and be enlightened in nose; and its prosperity is reflected in body hair. The Kidney phase should be generated in lung, govern bone, be enlightened in ears, and affect the appearance of hair. The Liver phase should be generated in kidney, govern tendon, be enlightened in eyes, and affect the appearance of “paw” (finger, toe, nail). Furthermore, Heart phase is connected with small intestine, Spleen phase is connected with stomach, Lung phase is connected with large intestine, Kidney phase is connected with bladder, and Liver phase is connected with gallbladder. The arrow along outer circle denotes the “mother-son” relationship, and the arrow along the diagonal direction indicates the “master-servant” relationship.
Heart phase includes insulin-like growth factor (IGF), angiotensin (Ang), and magnesium (Mg); Spleen phase includes atrial natriuretic peptide (ANP), aldosterone, retinoic acid and ghrelin; Lung phase includes fibroblast growth factor-7 (FGF7), vascular endothelial growth factor (VEGF), ascorbic acid (vitamin C) and hypoxia inducible factor [69]; Kidney phase includes calcitonin, parathyroid hormone-related protein (PTHrP), Wnt and nitric oxide (NO); Liver phase includes hepatocyte growth factor (HGF), erythropoietin (EPO), renin, superoxide dismutase [6], aldo-keto reductase [54] and glutathione [70].
Figure 2 The pathways of signaling molecules are consistent with the Five Phase relationships in TCM. The signaling molecules in each organ phase regulate each other according to Yang-Yi-Qi relationships in TCM. CTN: calcitonin; VC: ascorbic acid; VA: retinoic acid; ADO: aldosterone. Arrow means increasing, solid circle means decreasing, and arrow and circle together indicate regulating. The thick arrow with shallow color indicates the TCM relationship.
Figure 3 The popular TCM therapeutic strategies are consistent with the pathways of corresponding signaling molecules. Arrow means increasing, solid circle means decreasing, and arrow and circle together indicate regulating.
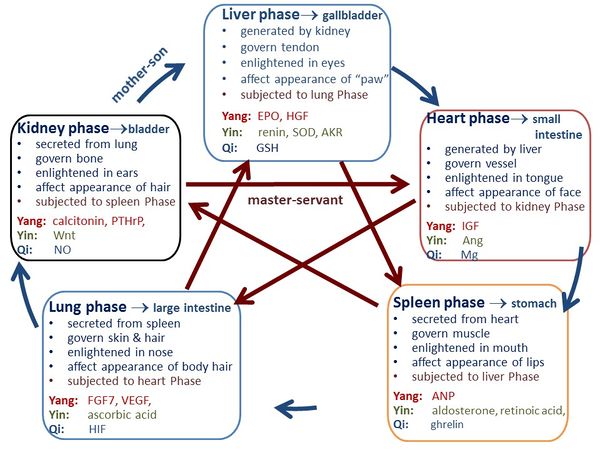

Figure 1. The functions and interrelationships of the Five Phases in TCM. The Heart phase should be generated in liver, govern blood vessel, and be enlightened in tongue; and its prosperity is reflected by face [2]. The Spleen phase should be generated in heart, govern muscle, and be enlightened in mouth; and its prosperity is reflected by lips. The Lung phase should be secreted in spleen, govern skin and hair, and be enlightened in nose; and its prosperity is reflected in body hair. The Kidney phase should be generated in lung, govern bone, be enlightened in ears, and affect the appearance of hair. The Liver phase should be generated in kidney, govern tendon, be enlightened in eyes, and affect the appearance of “paw” (finger, toe, nail). Furthermore, Heart phase is connected with small intestine, Spleen phase is connected with stomach, Lung phase is connected with large intestine, Kidney phase is connected with bladder, and Liver phase is connected with gallbladder. The arrow along outer circle denotes the “mother-son” relationship, and the arrow along the diagonal direction indicates the “master-servant” relationship.
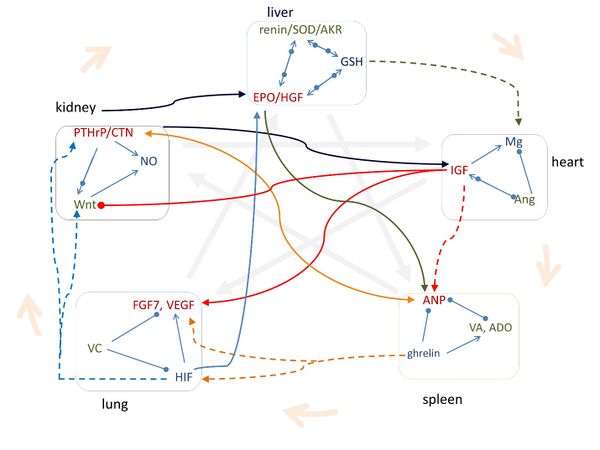

Figure 2. The pathways of signalling molecules are consistent with the Five Phase relationships in TCM. The signalling molecules in each organ phase regulate each other according to Yang-Yi-Qi relationships in TCM. CTN: calcitonin; VC: ascorbic acid; VA: retinoic acid; ADO: aldosterone. Arrow means increasing, solid circle means decreasing, and arrow and circle together indicate regulating. The thick arrow with shallow color indicates the TCM relationship.
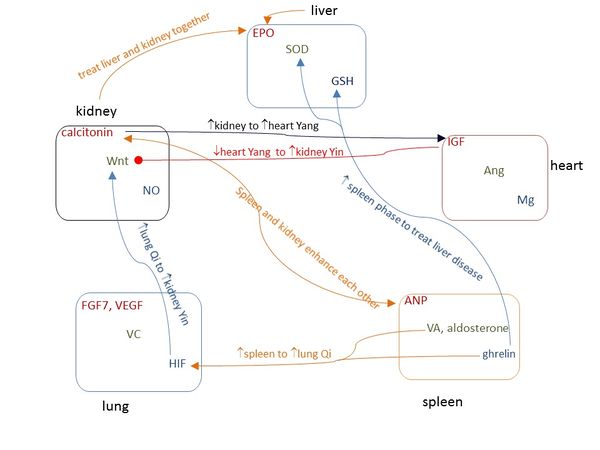

Figure 3. The popular TCM therapeutic strategies are consistent with the pathways of corresponding signalling molecules. Arrow means increasing, solid circle means decreasing, and arrow and circle together indicate regulating.
Supplementary Information
1. Identifying the signaling molecules for Yang, Yin and Qi in each organ phase of TCM
According to the “Yellow Emperor’s Inner Canon” (YEIC) [1], the fundamental doctrinal source for TCM, the Heart phase should be generated in liver organ, govern blood vessel, and be enlightened in tongue; and its prosperity is reflected by face [2]. The Spleen phase should be generated in heart organ, govern muscle, and be enlightened in mouth; and its prosperity is reflected by lips. The Lung phase should be secreted in spleen organ, govern skin and hair, and be enlightened in nose; and its prosperity is reflected in body hair. The Kidney phase should be generated in lung organ, govern bone, be enlightened in ears, and affect the appearance of hair. The Liver phase should be generated in kidney organ, govern tendon, be enlightened in eyes, and affect the appearance of “paw” (finger, toe, nail). Furthermore, Heart phase is connected with small intestine, Spleen phase is connected with stomach, Lung phase is connected with large intestine, Kidney phase is connected with bladder, and Liver phase is connected with gallbladder.
1.1 Key substances of Heart phase are IGF, Ang and Mg. According to YEIC [1], the Heart phase should exert significant effect on heart organ, be generated in liver, govern blood vessel, and be enlightened in tongue. Moreover, the prosperity of Heart phase is reflected by face [2].
In physiological reality, IGF is secreted by liver and is important to heart function. IGF-1 signaling regulates metabolism, contractility, senescence, autophagy, hypertrophy, and apoptosis in the heart; it activates canonical and noncanonical signaling pathways in the heart; deficiency in IGF-1 may drive cardiovascular disease, and local IGF-1 therapy can prevent heart injuries in experimental models [71]. IGF system exerts multiple physiologic effects on the vasculature, and plays a role in multiple vascular pathologies, including hypertension, diabetic vascular disease, atherosclerosis, angiogenesis, and restenosis [72]. Chronic treatment with IGF-1 significantly improves the vasoreactivity [73]. IGF is also enlightened in tongue, its level affects the tongue size, with increasing IGF level, tongue weights significantly increase with histopathological changes present; when IGF level is back to normal, tongue size returns to control levels [74]. Moreover, IGF level is reflected in skin, and high levels of IGF associate with reduced skin wrinkling [75]. As IGF-1 has warming effect [76], we identify IGF as Heart Yang.
Ang is secreted by liver; Ang II regulates cardiac and blood vessel contractility, and is involved in cardiac growth, remodeling, and apoptosis [77]. Angiotensin-converting enzyme inhibitor induces tongue angioedema, indicating an important role of Ang in tongue [78]. Moreover, the complete renin-angiotensin system is present in human skin and plays a role in normal cutaneous homeostasis [79]. As Ang II induces hypothermia [80], we identify Ang as Heart Yin.
Mg is important to normal heart rhythm and is a cofactor in more than 300 enzyme systems that regulate diverse biochemical reactions in the body, including blood pressure regulation and blood glucose control [81]. Mg relaxes blood vessel [82]; and it is also enlightened in tongue, as patients with burning mouth syndrome at the anterior tongue exhibit Mg deficiency as manifested by lower than normal Mg levels in saliva and erythrocytes [83]. Moreover, Mg affects the appearance of skin, it was found that magnesium-rich Dead Sea salt solution enhances skin hydration, improves skin barrier function, and reduces inflammation in atopic dry skin [84]. Due to its strong regulation and actuation effect, we identify Mg as Heart Qi.
TCM considers that Yang and Yin regulate each other and Figure 1 shows the interrelationships of IGF, Ang and Mg. It was found that Ang II infusion in rat decreases levels of circulating and skeletal muscle IGF-1 [85], Ang II stimulates cardiac IGF-1 [86] and transcription of IGF-1 receptor in vascular smooth muscle cells [87], and IGF-1 reduces Ang II [88]. Moreover, the exposure of the platelets to Ang II significantly decreases intracellular Mg concentrations [69], and Mg levels are strongly associated with IGF-1 [89].
1.2 Key substances of Spleen phase are ANP, aldosterone, retinoic acid, and ghrelin. According to YEIC [1], the Spleen phase should exert significant effect on spleen organ, be generated in heart, govern muscle, and be enlightened in mouth. Moreover, the prosperity of Spleen phase is reflected by lips.
In physiological reality, ANP is secreted by heart muscle cells that intervenes in the short- and long-term control of blood pressure and of water and electrolyte balance [90]. The spleen is an important site of ANP-induced fluid extravasation into the systemic lymphatic system. ANP enhances the extravasation of isoncotic fluid from the splenic vasculature both by raising intrasplenic microvascular pressure and by increasing filtration area [91]. ANP inhibits proliferation and hypertrophy of vascular smooth muscle cells and endothelial cells through cGMP production [92] and plays a physiological role on the modulation of salivary secretion [93]. It has been known that the sebum and color of lips of women are dependent on estrogen [94] and the feature of lips of men is dependent on testosterone [95], moreover, estrogen receptors increase ANP [96] and ANP induces testosterone production [97], indicating that lips reflect the level of ANP. As the famous function of Spleen Yang in TCM is “warming” and maintaining body temperature is an important function of ANP [98], we identify ANP as Spleen Yang.
Aldosterone retains needed salt in human body and helps control blood pressure, the balance of electrolytes in blood, and the distribution of fluids in the body. An intracardiac production of aldosterone was recently found in rat [99]. Aldosterone increases spleen size and weight [100], and stimulates vascular smooth muscle cell proliferation [101]. Aldosterone is enlightened in mouth, as low level of aldosterone causes dry mouth [102] and aldosterone refreshes mouth [103]. Moreover, estrogen receptor-β ligands modulate aldosterone synthesis [104] and short-term testosterone administration decreases aldosterone levels of men [105], indicating the appearance of lips reflects aldosterone level. Retinoic acid (vitamin A) controls the homeostasis of splenic cells [106] and is generated by the epicardium [107]. It also induces differentiation in both adult skeletal muscle primary satellite cells and a myogenic cell line [108], regulates oral epithelial differentiation [109], and serves as an effective treatment for oral leukoplakia [110]. Moreover, excess exogenous retinoic acid can induce cleft palate [111], indicating it affects the appearance of lips. Since either aldosterone or retinoic acid inhibits heat generation in body [112] [113], we identify them as Spleen Yin.
Ghrelin is a gastric peptide that regulates the distribution and consumption of energy. Ghrelin inhibits proliferation of splenic T cells [114] and modulates cardiac, smooth, and skeletal muscle functions [115]. Ghrelin produced in the oral cavity appears to play a regulatory role in innate immune responses to inflammatory infection [116]. Moreover, estrogen replacement therapy increased active plasma ghrelin levels [117] and testosterone correlates positively with ghrelin levels in men [118], indicating that ghrelin is reflected by the appearance of lips. In TCM, Spleen Qi has the function of delivering and transforming food energy; as ghrelin effectively regulates food intake and energy homeostasis [119], we identify it as Spleen Qi.
Figure 1 shows the contrary effect and supportive relationship between ANP and aldosterone/retinoic acid. ANP has exactly opposite function of aldosterone, ANP decreases circulating aldosterone and vice versa [120]. Retinoic acid signaling markedly stimulates natriuretic peptide receptor-A gene expression [121]. ANP inhibits proliferation of vascular smooth muscle [92], aldosterone stimulates muscle cell proliferation [101] and retinoic acid induces muscle differentiation [108]. Moreover, it was found that ANP level decreases significantly after treatment with ghrelin in a rat model [122]; ghrelin elevates aldosterone [123]; and applying ghrelin supplementation to normal lungs increased retinoic acid receptor α/γ expression [124].
1.3 Key substances of Lung phase are FGF7, VEGF, ascorbic acid and HIF. According to YEIC [1], these signaling molecules should exert significant effect on lung organ, be secreted in spleen, govern skin and hair, and be enlightened in nose. Moreover, the prosperity of Lung phase is reflected in body hair.
FGF7 is secreted by spleen [125] and is a potent mitogen that enhances cell proliferation in various organs, including the lung, skin, intestine, breast, and liver. It controls the lung morphogenesis, respiratory epithelial cell differentiation and proliferation [126]. FGF7-treated skin is significantly stronger than control vector-transfected skin, indicating that FGF7 can increase epithelial thickness and strength [127]. FGF7 is expressed in the hair follicle and may have several functions in the hair growth cycle [128]. FGF7 might play an important role in nasal epithelial proliferation and that excessive synthesis of FGF7 in nasal polyp stroma may contribute to hypertrophy of the nasal mucosa in patients with chronic sinusitis associated with nasal polyposis [129]. Due to its proliferation effect, we identify FGF7 as Lung Yang.
VEGF is critical for the development and maintenance of the lung and also plays a role in several acute and chronic lung diseases [130]. It was found that splenic T cells produce VEGF [131]. VEGF is the major mediator of hair follicle growth and cycling that promotes hair growth and increases hair follicle and hair size [132]. VEGF is also the major skin angiogenesis factor [133]. Moreover, VEGF promotes nasal epithelial cell growth and inhibits apoptosis [134]. The platelet-rich plasma injection has been used in the treatment of androgenetic alopecia, it was found that activated platelet-rich plasma prolongs anagen phase of hair growth cycle through increasing FGF7 level and increases the perifollicular vascular plexus through increasing VEGF level [135], indicating that both FGF7 and VEGF affect the appearance of hair. As the function of brown adipose tissue is to generate body heat and VEGF is important for brown adipose tissue development and maintenance [136], we identify VEGF as Lung Yang.
Ascorbic acid (Vitamin C) was identified in the early 1900s in the search for a deficient substance responsible for scurvy which was directly linked to pneumonia. Vitamin C is a physiological antioxidant protecting host cells against oxidative stress caused by infections, and plays a role on preventing and treating pneumonia [137] and other lung disease [138]. It was found that vitamin C content in spleen is much higher than that in other organs [139] and there exists an extracellular pool of ascorbic acid in lung maintained even during scurvy [140]. Dihydrotestosterone (DHT)-dependence causes hair loss, it was found that L-ascorbic acid 2-phosphate, a stable form of vitamin C, represses DHT-induced DKK-1 expression in cultured dermal papilla cells of human hair follicles [141]. Vitamin C is an essential component in producing collagen in body and a potent antioxidant that can help rejuvenate aged and photodamaged skin [142]. Ascorbic acid is also enlightened in nose, as Vitamin C is safe and natural therapies used to treat perennial allergic rhinitis [143]. Moreover, deficiency of ascorbic acid causes enlargement and keratosis of hair follicles mainly of the upper arms [144], indicating that the body hairs reflect the level of ascorbic acid. TCM considers that Lung Yin has “moisturizing lung” effect, therefore we identify ascorbic acid as Lung Yin.
HIF which is a highly conserved transcription factor that is present in almost all cell types, is tightly regulated by oxygen availability, and regulates the expression of hundreds of genes. HIF system plays a critical role in pulmonary development [145]. HIF-2α and HIF-1β are highly expressed in the precortex above the hair matrix, an area containing differentiating stem cells; and HIF-2α / HIF-1β complex regulates hair follicle differentiation [146]. Moreover, HIF-1α plays a crucial role in skin homeostasis and stress response [147] and HIF-1 mediates nasal polypogenesis [148]. Due to its strong regulation function, we identify HIF as Lung Qi.
Figure 1 shows the contrary and regulating interrelationship of HIF, FGF7, VEGF and vitamin C. HIF-1 activates VEGF transcription in hypoxic cells [149]. HIF promotes the expression of FGF7 mRNA levels [150]. Vitamin C prevents endothelial VEGF and VEGFR-2 overexpression [151] and inhibits NO-induced HIF-1α stabilization and accumulation [152]. Ascorbic acid stimulates collagen synthesis, but FGF inhibits collagen synthesis in the gels [153].
1.4 Key substances of Kidney phase are calcitonin, PTHrP, Wnt and NO. According to YEIC, Kidney phase should exert significant effect on kidney organ, be generated in lung, govern bone, and be enlightened in ears. Moreover, the prosperity of Kidney phase is reflected in hair.
Both calcitonin and PTHrP are secreted from lung [154] [155]. Calcitonin augments the renal reabsorptive capacity for calcium [156], enhances osteoblastic bone formation, and inhibits bone-resorbing activity of osteoclasts [157]. Calcitonin treatment can stabilize hearing loss or improve hearing in Paget's disease [158]. Moreover, calcitonin gene-related peptide (CGRP) treatment reduces the hair follicle pigmentation [159], indicating the appearance of hair reflects CGRP level. PTHrP is potent renal regulating factor and mitogenic for various renal cell types, and plays a role in renal development [160]. It plays a central role in the physiological regulation of bone formation by promoting recruitment and survival of osteoblasts, and probably plays a role in regulating bone resorption by enhancing osteoclast formation [161]. Moreover, PTHrP mRNA can be detected in the otic vesicle [162]. Furthermore, PTHrP may induce premature catagen-telogen transition in hair growth [163], indicating that PTHrP may affect the appearance of hair. TCM considers that the important function of Kidney Yang is “warming”, as both calcitonin and PTHrP have “warming” effect [164]; [165], we identify both calcitonin and PTHrP as Kidney Yang.
Wnt signaling regulates cell-to-cell interactions during development and adult tissue homeostasis, and is associated with kidney development and kidney diseases [166]. Several Wnt genes are expressed in lung [167]. Wnt signaling has potential roles in bone remodeling in both physiological and pathological conditions, it acts on osteoblast precursor cells and promote their differentiation into mature osteoblasts, and it can either suppress bone resorption or enhance osteoclast formation through various signaling pathway [168]. Wnt is also enlightened in ears, as Wnt signaling is active in dorsal regions of the otic vesicle, where it functions to regulate the expression of genes (Dlx5/6 and Gbx2) necessary for vestibular morphogenesis [169]. Wnt signaling is a key pathway that initiates pigmented hair regeneration [170], indicating the appearance of hair reflects Wnt level. Since Wnt signaling blocks thermogenesis [171], we identify Wnt as Kidney Yin.
NO is the second messenger and a major regulator in the cardiovascular, immune, and nervous systems. In the kidney NO has numerous important functions including the regulation of renal hemodynamics, maintenance of medullary perfusion, mediation of pressure-natriuresis, blunting of tubuloglomerular feedback, inhibition of tubular sodium reabsorption, and modulation of renal sympathetic neural activity [172]. It is found that both osteoclasts and osteoblasts have inducible NO synthase and produce NO, whereas NO inhibits the proliferation of osteoclasts and decreases the proliferation of osteoblasts [173]. NO also plays an important role in the inner ear to regulate physiological reactions in both cochlea and vestibular parts of the labyrinth [174]. Moreover, NO regulates hair follicle activity [175], indicating that NO would affect the appearance of hair. Due to its strong regulation function, we identify NO as kidney Qi.
Figure 1 shows the interrelationships among calcitonin, PTHrP, Wnt and NO. Calcitonin stimulates expression of sclerostin [176] which is an inhibitor of Wnt, and administration of PTHrP can increase Wnt signaling [177]. Moreover, calcitonin increases plasma NO levels [178], PTHrP activates NO release [179], and Wnt-5a increases NO production [180]. Calcitonin enhances osteoblastic bone formation and inhibits bone-resorbing activity of osteoclasts [157]. PTHrP promotes recruitment and survival of osteoblasts and enhances osteoclast formation [161]. Wnt signaling stimulates osteoblastogenesis [181] and decreases osteoclast formation [182].
1.5 Key substances of Liver phase are EPO, HGF, renin, SOD, AKR, and GSH. According to YEIC, Liver phase should exert significant effect on liver organ, be generated in kidney, govern tendon, and be enlightened in eyes. Moreover, the prosperity of Liver phase is reflected in “paw” (finger, toe, nail).
EPO is a glycoprotein that promotes the proliferation and differentiation of erythrocyte precursors. It was reported that high-dose EPO increases liver regeneration by affecting the biochemical, morphological, and histopathological parameters after liver resection [183]. EPO is produced principally in the liver during fetal gestation and mainly in the kidney for adult [184]. It is used to provide protective and regenerative activity in non-haematopoietic tissues and stimulates tendon healing [185]. EPO is a potent angiogenic factor observed in the diabetic human and mouse eyes [186]. Moreover, digital clubbing is defined by structural changes at the base of the nails which is associated with increased EPO levels [187], and patients using erythropoiesis stimulating agents have lower rate of onychomycosis compared to those not using them [188], indicating that the appearance of finger reflects the EPO level. Since EPO increases brown fat gene expression [189] and has proliferation effect, we identify EPO as Liver Yang.
YEIC also denotes that Kidney Phase produces bone, and bone produces Liver Phase [1]. HGF is produced by mesenchymal cells, and the bone marrow stem contains mesenchymal stem cells. HGF accelerates both the hepatic regeneration and function in vivo [190]. Platelet-rich plasma containing HGF is widely used in orthopaedic/sports medicine to repair injured tendons [191] and local administration of recombinant HGF promotes the adhesive healing process at the tendon-bone junction both histologically and mechanically [192]. HGF is also enlightened in eyes. There is growing evidence that HGF is synthesized by autocrine loops in a number of cell systems prominent among which are ocular cells such as the corneal endothelium, the lens epithelium, the retinal pigment epithelium and others; furthermore, HGF is a constituent of tears, aqueous humour and vitreous humour [193]. Moreover, digital clubbing is stimulated by high level of HGF [194], indicating that the appearance of fingers reflects the HGF level. As HGF plays a central role in adipose tissue development [195] and has proliferation effect, we identify HGF as Liver Yang.
Renin is produced in kidney and exerts significant effect on liver. The renin-substrate concentration in patients with liver disease is much lower than that of normal subjects [196]. Loading-induced periodontal ligament tissue homeostasis is regulated through the renin–angiotensin system [197]. Renin is also present in human eyes [198]. Moreover, the psoriasis usually affects the fingernails and toenails, causing pitting, thickening, and irregular contour of the nail; it was found that plasma renin activity was enhanced in psoriatic patients [199], indicating that renin would affect the appearance of nails. As blockade of the renin-angiotensin system improves liver regeneration [200], we identify renin as Liver Yin.
SOD is an important antioxidant defense in nearly all living cells exposed to oxygen and is intensively expressed in kidney [201]. SOD mimetic improves the function, growth and survival of small size liver grafts after transplantation in rats [202] and Cu/Zn SOD is broadly distributed in liver [203]. Mn-SOD is excessively expressed in the cultured fibroblasts derived from the stress-deprived patellar tendon in the rat [204]. SOD is also enlightened in eyes. The retina contained much CuZn-SOD activity and the cornea and sclera contained several times more SOD activity than the lens [205]. Moreover, the levels of MnSOD are reduced in psoriasis [206], indicating that SOD would affect the appearance of nails. It was found that cold-exposure activates brown adipose tissue thermogenesis but decreases SOD expression [207], indicating SOD has “cooling” effect, therefore we identify SOD as Liver Yin.
AKR is expressed in kidney [208]. The AKR superfamily comprises of several enzymes that catalyze redox transformations involved in intermediary metabolism, detoxification and biosynthesis [209]. AKR-7A protects liver cells and tissues from acetaminophen-induced oxidative stress and hepatotoxicity [210]. It was found that the AKR content is much higher in patients with tendon xanthomas, indicating that AKR plays important role in tendon [211]. AKR is also enlightened in eyes, as expression of both AKR1B1 and AKR1B10 can be observed in cornea, iris, ciliary body, lens, and retina [212]. Cold-exposure suppresses AKR level significantly [213], indicating AKR has “cooling” effect, therefore we identify AKR as Liver Yin.
GSH is a substance produced naturally by the liver and effectively scavenges free radicals and other reactive oxygen species [214]. GSH plays a key role in the liver in detoxification reactions and has close relationship with tendon and eyes. It was reported that GSH level was significantly decreased in post-injury chicken models and increasing GSH can reduce the extent of adhesion of healing tendons [215] . GSH and the related enzymes belong to the defense system protecting the eye against chemical and oxidative stress; ageing, cataract, diabetes, irradiation and administration of some drugs could cause changes of GSH and related enzymes activity in lens, cornea, retina and other eye tissues [216]. Moreover, nail hyperpigmentation involves decreased GSH levels [217], indicating the appearance of nail reflects the GSH level. In TCM, Liver Qi has “catharsis” function; due to its strong detoxification effect, we identify GSH as Liver Qi.
Figure 1 shows the interrelationships among EPO, HGF, renin, SOD, AKR and GSH. HGF raises cellular GSH levels as well as SOD1 expression [218] [219], and downregulates aldose reductase [220] which is the prototypical enzyme of the AKR superfamily [221]. When treated with EPO, the GSH and SOD levels in kidney of rats are significantly decreased [222]. The extracellular SOD is a major repressor of hypoxia-induced EPO gene expression [223], but treatment with EPO increases vascular expression of SOD1 [224]. SOD and GSH work together to prevent or repair the damage caused by reactive oxygen species. SODs convert superoxide radical into hydrogen peroxide and molecular oxygen, while the GSH peroxidases (GPx) convert hydrogen peroxide into water, and GSH helps the productivity of GPx [225]. GSH and AKR are also interrelated, since GSH levels are reduced in AKR-deficient strains [226]. Furthermore, renin-angiotensin system stimulates EPO secretion [227], blockade of the renin-angiotensin system enhances hepatic HGF production [228], and inhibition of GSH increases plasma renin activity [229].
2. Effects of TCM herbal medicines on signaling molecules
TCM herbal medicines are categorized into various functions to regulate Yang-Yin-Qi in each organ. We now validate our results by exploring whether the Chinese herbal medicine can regulate the corresponding signaling molecules. Table S1 lists some representative herbal medicines and their function on the corresponding signaling molecules.
Ginseng is a famous TCM herbal medicine which can strengthen Spleen phase particularly in enhancing Spleen Qi and benefit Lung phase particularly in augmenting Lung Qi [230]. In physiological reality, ginseng increases ANP level [231] and enhances the ability of all-trans retinoic acid (ATRA) to induce terminal differentiation of leukemia cell [232]; ginsenoside Rb1 significantly stimulates the ghrelin endocrine to inhibit endothelial injury [233]. Ginsenoside-Rg1 effectively relieves aldosterone-induced oxidative stress, thereby indirectly inhibiting aldosterone-induced podocyte autophagy [234]. In human keratinocyte cultures, ginsenoside Rb1 enhances VEGF production and significantly increases expression of HIF-1α [235].
Astragalus is a popular TCM herbal medicine used to enhance Lung Qi and Lung Yang and increase Spleen Yang [230]. In physiological reality, flavonoids from radix astragali increase the expression of HIF-1α in both mRNA and protein markedly [236] and enhance VEGF mRNA expression [237]. Furthermore, astragalus induces the natriuresis by significantly improving the renal reaction to ANP [238].
Deer antler is a famous Chinese herbal medicine for strengthening Kidney Yang and benefiting blood, and it principally affects Kidney and Liver phases [230]. In physiological reality, deer antler contains plenty of calcitonin and PTHrP, calcitonin content increases significantly during the rapid antler growth period [239], and intense and overlapping expression of PTHrP and its receptor exists in developing osseocartilaginous structures and in the underlying layers of velvet epidermis of deer antler [240]. Furthermore, deer antler increases serum EPO value [241], SOD activity in serum [242], and glutathione peroxidase [243], which is consistent with that deer antler benefits Liver phase.
In TCM, cynomorium songaricum has the function to warm Kidney Yang and nourish the Liver Yin [244]. In physiological reality, cynomorium songaricum significantly increases calcitonin [245] and increases CuZn-SOD and glutathione peroxidase [246].
Angelica sinensis is famous for enhancing hematopoiesis, and it principally affects Liver and Heart phases [230]. In physiological reality, angelica stimulates EPO to enhance hematopoiesis [247]; [248], promotes HGF [249], increases GSH [250], enhances the activity of SOD [251], and regulates renin activities [252]. Moreover, angelica sinensis polysaccharides stimulates IGF-1 expression [253] but inhibits Ang II [252].
In TCM, cordyceps sinensis is famous to moderately enhance Lung phase [230]. In physiological reality, cordyceps sinensis up-regulates the expression of HIF-1α [254], maintains higher HIF-1 level in human lung epithelial cells [255], contains higher ascorbic acid contents [256], and lowers VEGF level in lung [257].
In TCM, Lilium lancifoliium is famous to nourish Lung Yin [230]. In China, Lilium lancifoliium is used as an edible lily and is an important source of ascorbic acid [258].
In TCM, wolfberry (lyceum barbarum) is famous to nourish Kidney Yin and Liver Yin [230]. In physiological reality, wolfberry phytochemicals activate Wnt signaling [259], and increase SOD and GPx in blood [260].
TCM formula is a combination of herbs with suitable dosage to treat symptoms, and some formulas are particularly used to regulate Yang-Yin-Qi of organs. Table S2 lists some representative TCM formulas and shows that these TCM formulas regulate the corresponding signaling molecules. Liujunzi Tang (Six Gentlemen Decoction, consisting of ginseng, bighead atractylodes rhizome, poria, roasted licorice root, tangerine peel and pinellia tuber) is used to replenish Spleen Qi [261], it is widely used for the treatment of the upper gastrointestinal symptoms of patients with functional dyspepsia, gastroesophageal reflux disease, dyspeptic symptoms of postgastrointestinal surgery patients, and chemotherapy-induced dyspepsia in cancer patients, and oral administration of Liujunzi Tang potentiates orexigenic action of ghrelin [262].
Yiguan Decoction (consisting of glehnia root, ophiopogon root, Chinese angelica root, dried rehmannia root, wolfberry, Sichuan chinaberry) is used to nourish Liver Yin and sooth Liver Qi in TCM [261]. It was found that Yiguan Decoction significantly increases protein expressions of Cu/Zn SOD, glutathione S-transferase (GST) and AKR family 7, while notably decrease the expression of glutathione synthetase (GSS) [263].
Liuwei Dihuang Wan (Rehmannia Bolus with Six Herbs, consisting of prepared rhizome of rehmannia, dogwood fruit, dried Chinese yam, oriental water plantain, poria, moutan bark) is used to nourish and enrich Kidney Yin in TCM [261]. In physiological reality, it could up-regulate canonical Wnt/beta-catenin signaling [264].
Zuogui Pill (Back to Left Pill, consisting of prepared rhizome of rehmannia, the seed of Chinese dodder, achyranthes bidentate, glue of tortoise plastron, deerhorn glue, Chinese yam, dogwood fruit, and wolfberry) is used to nourish Kidney Yin in TCM [265], and it was found that Zuogui Pill increases Wnt1 in Wnt signal transduction pathway [266].
ShenQi Pill (Kidney Qi Pill, consisting of dried rehmannia root, Chinese yam, dogwood fruit, oriental water plantain, poria, moutan bark, cinnamon twig, and prepared aconite root) is used to enhance Kidney Qi [261]. In physiological reality, it can significantly increase NO production [267].
Zonghai Tang [1851-1908] was a famous Chinese physician, he stated that “replenishing Spleen Yin is for appetite” [50]; to replenish spleen Yin, he suggested to use “Mai-men-dong Tang” (Ophiopogon Japonicus Decoction) formulated by Jing-Yue Zhang (1563-1640) [8]. Ophiopogon Japonicus Decoction consists of ophiopogon japonicus and japonica rice, in which ophiopogon japonicus contains vitamin A [230]. In China, japonica rice grows up in temperate region, it was found that temperate japonica rice has the highest levels of carotenoids among the types of rice [268], and carotenoids are a group of plant pigments important in the human diet as the only precursors of vitamin A [269].
Table S1.
Consistence between effects of Chinese herbal medicine on TCM functions and on signaling molecules.
| herbal medicine | TCM functions | effect on signaling molecules |
| ginseng | Spleen phase [230]
|
ANP[231], ATRA[232],
ghrelin[233], aldosterone[234] |
| Lung phase[230] | VEGF, HIF [235] | |
| astragalus | Lung Qi,
Lung Yang [230] |
HIF [236]
VEGF [237] |
| Spleen Yang [230] | ANP [238] | |
| deer antler | Kidney Yang [230] | contain calcitonin & PTHrP [239, 240] |
| benefit blood,
affect Liver phase [230] |
EPO[241], SOD[242],
GSH px[243] | |
| cynomorium
songaricum |
warm Kidney Yang [244] | calcitonin [245] |
| nourish Liver Yin [244] | SOD, GSH px [246] | |
| angelica sinensis | hematopoiesis,
affect LIVER |
EPO[247, 248], HGF[249], GSH[250], SOD[251], renin[252] |
| affect HEART [230] | IGF[253], Ang II [252] | |
| cordyceps sinensis
|
Lung phase[230] |
HIF[254, 255], ascorbic acid[256], VEGF[257] |
lilium lancifoliium |
nourish Lung Yin[230] |
Contain ascorbic acid[258] |
wolfberry |
Kidney Yin and Liver Yin[230] |
Wnt[259],SOD, GSH px[260] |
Table S2.
Consistence between effects of TCM formula on TCM functions and on signaling molecules.
TCM formula |
TCM functions |
effect on signaling molecules |
Liujunzi Tang (Six Gentlemen Decoction) |
Spleen Qi [261] |
ghrelin [262] |
Yiguan Decoction |
Liver Yin |
SOD, AKR [263] |
| sooth Liver Qi [261] | GST, GSS [263] | |
| Liuwei Dihuang Wan (Rehmannia Bolus with Six Herbs) | Kidney Yin [261] | Wnt [264] |
| Zuogui Pill (Back to Left Pill) | Kidney Yin[265] | Wnt [266] |
| Shenqi Pill (Kidney Qi Pill) | Kidney Qi [261] | NO [267] |
| Mai-Men-Dong Tang (Ophiopogon Japonicus Decoction) | spleen Yin [50] | vitamin A [230, 268, 269] |
References
1. Huang-Di, Huang Di Nei Jing [Yellow Emperor's Inner Canon]. . 2005, Beijing: People's Health Publishing House
2. Parrott, J.A., et al., Autocrine interactions of keratinocyte growth factor, hepatocyte growth factor, and kit-ligand in the regulation of normal ovarian surface epithelial cells. Endocrinology, 2000. 141(7): p. 2532-2539.
3. YE81, Huang Di 81 Nan Jing [Yellow Emperor's Classic on 81 Medical Problems]. . 1996, ShenYang, Liaoning, China: Liaoning Education Press.
4. Liu, Y., et al., Association of serum retinoic acid with hepatic steatosis and liver injury in nonalcoholic fatty liver disease. Am J Clin Nutr, 2015. 102(1): p. 130-7.
5. Cao, Y.K., et al., Cardioprotective Effect of Ghrelin in Cardiopulmonary Bypass Involves a Reduction in Inflammatory Response. Plos One, 2013. 8(1).
6. Zwirska-Korczala, K., et al., Role of leptin, ghrelin, angiotensin II and orexins in 3T3 L1 preadipocyte cells proliferation and oxidative metabolism. J Physiol Pharmacol, 2007. 58 Suppl 1: p. 53-64.
7. Porat, O., et al., Erythropoietin stimulates atrial natriuretic peptide secretion from adult rat cardiac atrium. J Pharmacol Exp Ther, 1996. 276(3): p. 1162-8.
8. Zhang, J., Jing Yue Quan Shu [Zhang Jing-Yue's Medical Book]. 1999, Beijing: China Press of Traditional Chinese Medicine
9. Vesely, D.L., et al., Atrial natriuretic peptides increase calcitonin gene-related peptide within human circulation. Metabolism-Clinical and Experimental, 1997. 46(7): p. 818-825.
10. Yamamoto, A., et al., Calcitonin Gene-Related Peptide (Cgrp) Stimulates the Release of Atrial Natriuretic Peptide(Anp) from Isolated Rat Atria. Biochemical and Biophysical Research Communications, 1988. 155(3): p. 1452-1458.
11. Zhou, Z., Zhou ShenZhai Yixue Quanshu [Zhou ShenZhai Medical Book]. . 2010, Hainan, China: Hainan Publishing House
12. Farley, J., et al., Calcitonin increases the concentration of insulin-like growth factors in serum-free cultures of human osteoblast-line cells. Calcified Tissue International, 2000. 67(3): p. 247-254.
13. Schlupf, J. and H. Steinbeisser, IGF antagonizes the Wnt/beta-Catenin pathway and promotes differentiation of extra-embryonic endoderm. Differentiation, 2014. 87(5): p. 209-219.
14. Landon, R.A. and E.A. Young, Role of magnesium in regulation of lung function. J Am Diet Assoc, 1993. 93(6): p. 674-7.
15. Hung, C.F., et al., Role of IGF-1 pathway in lung fibroblast activation. Respiratory Research, 2013. 14.
16. Slomiany, M.G. and S.A. Rosenzweig, IGF-1-induced VEGF and IGFBP-3 secretion correlates with increased HIF-1 alpha expression and activity in retinal pigment epithelial cell line D407. Investigative Ophthalmology & Visual Science, 2004. 45(8): p. 2838-2847.
17. Pereira, C.T., et al., Liposomal gene transfer of keratinocyte growth factor improves wound healing by altering growth factor and collagen expression. Journal of Surgical Research, 2007. 139(2): p. 222-228.
18. Jeschke, M.G. and D.N. Herndon, The combination of IGF-I and KGF cDNA improves dermal and epidermal regeneration by increased VEGF expression and neovascularization. Gene Therapy, 2007. 14(16): p. 1235-1242.
19. Stirling, D., et al., Angiotensin-Ii Inhibits Luteinizing Hormone-Stimulated Cholesterol Side-Chain Cleavage Expression and Stimulates Basic Fibroblast Growth-Factor Expression in Bovine Luteal Cells in Primary Culture. Journal of Biological Chemistry, 1990. 265(1): p. 5-8.
20. Pupilli, C., et al., Angiotensin II stimulates the synthesis and secretion of vascular permeability factor vascular endothelial growth factor in human mesangial cells. Journal of the American Society of Nephrology, 1999. 10(2): p. 245-255.
21. Takase, H.M., et al., FGF7 is a functional niche signal required for stimulation of adult liver progenitor cells that support liver regeneration. Genes & Development, 2013. 27(2): p. 169-181.
22. Taniguchi, E., et al., Expression and role of vascular endothelial growth factor in liver regeneration after partial hepatectomy in rats. Journal of Histochemistry & Cytochemistry, 2001. 49(1): p. 121-129.
23. Ipsen, D.H., P. Tveden-Nyborg, and J. Lykkesfeldt, Does Vitamin C Deficiency Promote Fatty Liver Disease Development? Nutrients, 2014. 6(12): p. 5473-5499.
24. Ersoz, G., et al., Management of fatty liver disease with vitamin E and C compared to ursodeoxycholic acid treatment. Turk J Gastroenterol, 2005. 16(3): p. 124-8.
25. Nath, B. and G. Szabo, Hypoxia and hypoxia inducible factors: Diverse roles in liver diseases. Hepatology, 2012. 55(2): p. 622-633.
26. Westenbrink, B.D., et al., Vascular endothelial growth factor is crucial for erythropoietin-induced improvement of cardiac function in heart failure. Cardiovascular Research, 2010. 87(1): p. 30-39.
27. Johnston, C.S., C.G. Meyer, and J.C. Srilakshmi, Vitamin-C Elevates Red-Blood-Cell Glutathione in Healthy-Adults. American Journal of Clinical Nutrition, 1993. 58(1): p. 103-105.
28. Wang, G.L., et al., Hypoxia-Inducible Factor-1 Is a Basic-Helix-Loop-Helix-Pas Heterodimer Regulated by Cellular O-2 Tension. Proceedings of the National Academy of Sciences of the United States of America, 1995. 92(12): p. 5510-5514.
29. Tajima, M., et al., The redox state of glutathione regulates the hypoxic induction of HIF-1. European Journal of Pharmacology, 2009. 606(1-3): p. 45-49.
30. Li ZZ, G.Q., Yi Zong Bi Du [Required Readings for Medical Professionals], . 1998, Beijing: People's Health Publishing House.
31. Obara, N., et al., Repression via the GATA box is essential for tissue-specific erythropoietin gene expression. Blood, 2008. 111(10): p. 5223-5232.
32. Konturek, P.C., et al., Ghrelin-induced gastroprotection against ischemia-reperfusion injury involves an activation of sensory afferent nerves and hyperemia mediated by nitric oxide. European Journal of Pharmacology, 2006. 536(1-2): p. 171-181.
33. Fernandez-Martinez, A.B., M.I.A. Jimenez, and F.J.L. Cazana, Retinoic acid increases hypoxia-inducible factor-1 alpha through intracrine prostaglandin E-2 signaling in human renal proximal tubular cells HK-2. Biochimica Et Biophysica Acta-Molecular and Cell Biology of Lipids, 2012. 1821(4): p. 672-683.
34. Mackenzie, I.C. and Z.R. Gao, Keratinocyte growth factor expression in human gingival fibroblasts and stimulation of in vitro gene expression by retinoic acid. Journal of Periodontology, 2001. 72(4): p. 445-453.
35. Mazumdar, J., et al., O-2 regulates stem cells through Wnt/beta-catenin signalling. Nature Cell Biology, 2010. 12(10): p. 1007-1013.
36. Pelosi, M., et al., Parathyroid hormone-related protein is induced by hypoxia and promotes expression of the differentiated phenotype of human articular chondrocytes. Clinical Science, 2013. 125(9-10): p. 461-470.
37. Agani, F.H., et al., Role of nitric oxide in the regulation of HIF-1 alpha expression during hypoxia. American Journal of Physiology-Cell Physiology, 2002. 283(1): p. C178-C186.
38. Zhu, X.J., et al., BMP-FGF signaling axis mediates Wnt-induced epidermal stratification in developing mammalian skin. PLoS Genet, 2014. 10(10): p. e1004687.
39. Nejak-Bowen, K.N., et al., beta-Catenin Regulates Vitamin C Biosynthesis and Cell Survival in Murine Liver. Journal of Biological Chemistry, 2009. 284(41): p. 28115-28127.
40. d'Uscio, L.V., et al., Long-term vitamin C treatment increases vascular tetrahydrobiopterin levels and nitric oxide synthase activity. Circulation Research, 2003. 92(1): p. 88-95.
41. Barbagallo, M., et al., Effects of glutathione on red blood cell intracellular magnesium - Relation to glucose metabolism. Hypertension, 1999. 34(1): p. 76-82.
42. Chu, C.H., et al., IGF-II/mannose-6-phosphate receptor signaling induced cell hypertrophy and atrial natriuretic peptide/BNP expression via G alpha q interaction and protein kinase C-alpha/CaMKII activation in H9c2 cardiomyoblast cells. Journal of Endocrinology, 2008. 197(2): p. 381-390.
43. Chao YF, D.G., Zhu Bing Yuan Hou Lun Jiao Zhu [Annotation on General Treatise on Causes and Manifestations of All Diseases]. Vol. 14 & 27. 1992, Beijing: People's Health Publishing House.
44. Yildiz, B., et al., Increased serum levels of insulin-like growth factor (IGF)-1 and IGF-binding protein-3 in Henoch-Schonlein purpura. Tohoku Journal of Experimental Medicine, 2008. 214(4): p. 333-340.
45. Tojo, A., et al., Spironolactone with ACE inhibitor is effective in gross hematuria caused by nephroptosis. International Journal of Urology, 2006. 13(7): p. 990-992.
46. Morrone, L.F., et al., Interference of angiotensin-converting enzyme inhibitors on erythropoiesis in kidney transplant recipients: role of growth factors and cytokines. Transplantation, 1997. 64(6): p. 913-8.
47. Li, D., Pi Wei Lun [Treatise on the spleen and stomach]. . 2007, Beijing: China Press of Traditional Chinese Medicine.
48. Gower, W.R., et al., Gastric atrial natriuretic peptide regulates endocrine secretion in antrum and fundus of human and rat stomach. American Journal of Physiology-Gastrointestinal and Liver Physiology, 2003. 284(4): p. G638-G645.
49. Inui, A., et al., Ghrelin, appetite, and gastric motility: the emerging role of the stomach as an endocrine organ. Faseb Journal, 2004. 18(3): p. 439-456.
50. Tang, R., Xue Zheng Lun [Blood Syndrome Theory]. 1977, Shanghai: Shanghai People's Publishing House
51. Li, S., Compendium of Materia Medica. . 2002, Beijing China Economic Publishing House.
52. Chung, Y.C., Y.C. Hou, and A.C.H. Pan, Endoglin (CD105) expression in the development of haemorrhoids. European Journal of Clinical Investigation, 2004. 34(2): p. 107-112.
53. Lee, W.T., et al., Increased prevalence of constipation in pre-school children is attributable to under-consumption of plant foods: A community-based study. J Paediatr Child Health, 2008. 44(4): p. 170-5.
54. Ghambarali, Z., et al., Ethanolic extract of Ficus carica leave Suppresses Angiogenesis by Regulating VEGF-A and Integrin beta3 mRNA Expression in Human umbilical vein endothelial cells. Indian J Physiol Pharmacol, 2014. 58(4): p. 407-15.
55. USDA, N.N.D. Statistics Report 09089, Figs, raw. Available from: https://ndb.nal.usda.gov/ndb/foods/show/2201?format=Stats&reportfmt=pdf...
56. Andersson, K.E. and K. Persson, Nitric-Oxide Synthase and Nitric Oxide-Mediated Effects in Lower Urinary-Tract Smooth Muscles. World Journal of Urology, 1994. 12(5): p. 274-280.
57. Balat, A., et al., Nitric oxide synthase gene polymorphisms in children with primary nocturnal enuresis: A preliminary study. Renal Failure, 2007. 29(1): p. 79-83.
58. Sudo, H., et al., Nicorandil, a Potassium Channel Opener and Nitric Oxide Donor, Improves the Frequent Urination without Changing the Blood Pressure in Rats with Partial Bladder Outlet Obstruction. Biological & Pharmaceutical Bulletin, 2008. 31(11): p. 2079-2082.
59. Sun, S., Zhong Yi Gan Dan Bing Liang Fang [Chinese medicine recipe for hepatology], . 2005, Beijing: JinDun Publishing House
60. Geetha, A., Evidence for oxidative stress in the gall bladder mucosa of gall stone patients. J Biochem Mol Biol Biophys, 2002. 6(6): p. 427-32.
61. Pomelov, V.S., et al., [Glutathione levels and the activity of the enzymes of glutathione metabolism in erythrocytes of patients with acute cholecystitis]. Sov Med, 1991(8): p. 27-30.
62. He R, T.R., Chen SS, Wu SK, Zhu ZT, Lu ZQ, Wei ZC, Zhang PQ, Chen ZL. , Gao Xue Ya Zheng Zhi [Hypertension Treatment]. . J Tradit Chin Med 1986. 2: p. 11-14.
63. Vaziri, N.D., Mechanism of erythropoietin-induced hypertension. American Journal of Kidney Diseases, 1999. 33(5): p. 821-828.
64. Morishita, R., et al., Hepatocyte Growth Factor (HGF) as a potential index of severity of hypertension. Hypertension Research-Clinical and Experimental, 1999. 22(3): p. 161-167.
65. Conn, J.W., D.R. Rovner, and E.L. Cohen, Suppression of Plasma Renin Activity in Primary Aldosteronism - Distinguishing Primary from Secondary Aldosteronism in Hypertensive Disease. Jama-Journal of the American Medical Association, 1964. 190(3): p. 213-&.
66. Nozik-Grayck, E., et al., Selective depletion of vascular EC-SOD augments chronic hypoxic pulmonary hypertension. American Journal of Physiology-Lung Cellular and Molecular Physiology, 2014. 307(11): p. L868-L876.
67. Tyther, R., et al., Protein carbonylation in kidney medulla of the spontaneously hypertensive rat. Proteomics Clinical Applications, 2009. 3(3): p. 338-346.
68. Cheng, P.W., et al., Wnt Signaling Regulates Blood Pressure by Downregulating a GSK-3 beta-Mediated Pathway to Enhance Insulin Signaling in the Central Nervous System. Diabetes, 2015. 64(10): p. 3413-3424.
69. Touyz, R.M. and E.L. Schiffrin, The Effect of Angiotensin-Ii on Platelet Intracellular Free Magnesium and Calcium Ionic Concentrations in Essential-Hypertension. Journal of Hypertension, 1993. 11(5): p. 551-558.
70. van Deel, E.D., et al., Extracellular superoxide dismutase protects the heart against oxidative stress and hypertrophy after myocardial infarction. Free Radical Biology and Medicine, 2008. 44(7): p. 1305-1313.
71. Troncoso, R., et al., New insights into IGF-1 signaling in the heart. Trends in Endocrinology and Metabolism, 2014. 25(3): p. 128-137.
72. Delafontaine, P., Y.H. Song, and Y.X. Li, Expression, regulation, and function of IGF-1, IGF-1R, and IGF-1 binding proteins in blood vessels. Arteriosclerosis Thrombosis and Vascular Biology, 2004. 24(3): p. 435-444.
73. Xu, Z.P., et al., Effects of insulin-like growth factor-1 on the relaxation responses of the cavernous smooth muscle from aged rats. Scandinavian Journal of Urology, 2015. 49(3): p. 260-266.
74. Kobayashi, A., et al., Morphological and histopathological changes in tongues of experimentally developed acromegaly-like rats. Hormone and Metabolic Research, 2006. 38(3): p. 146-151.
75. Noordam, R., et al., Serum insulin-like growth factor 1 and facial ageing: high levels associate with reduced skin wrinkling in a cross-sectional study. British Journal of Dermatology, 2013. 168(3): p. 533-538.
76. Sanchez-Alavez, M., et al., Insulin-like Growth Factor 1-mediated Hyperthermia Involves Anterior Hypothalamic Insulin Receptors. Journal of Biological Chemistry, 2011. 286(17): p. 14983-14990.
77. De Mello, W.C. and A.H.J. Danser, Angiotensin II and the heart - On the intracrine renin-angiotensin system. Hypertension, 2000. 35(6): p. 1183-1188.
78. Mlynarek, A., A. Hagr, and K. Kost, Angiotensin-converting enzyme inhibitor-induced unilateral tongue angioedema. Otolaryngology-Head and Neck Surgery, 2003. 129(5): p. 593-595.
79. Steckelings, U.M., et al., Human skin: source of and target organ for angiotensin II. Experimental Dermatology, 2004. 13(3): p. 148-154.
80. Wilson, K.M. and M.J. Fregly, Angiotensin Ii-Induced Hypothermia in Rats. Journal of Applied Physiology, 1985. 58(2): p. 534-543.
81. NIH-library-Mg. Magnesium. Available from: [https:// https://]www.nlm.nih.gov/medlineplus/druginfo/natural/998.html.
82. Dean, C., The magnesium miracle : discover the essential nutrient that will lower therisk of heart disease, prevent stroke and obesity, treat diabetes, and improve mood and memory. Ballantine books trade pbk. ed. 2007, New York: Ballantine Books. xxii, 309 p.
83. Henkin, R.I., V. Gouliouk, and A. Fordyce, Distinguishing patients with glossopyrosis from those with oropyrosis based upon clinical differences and differences in saliva and erythrocyte magnesium. Archives of Oral Biology, 2012. 57(2): p. 205-210.
84. Proksch, E., et al., Bathing in a magnesium-rich Dead Sea salt solution improves skin barrier function, enhances skin hydration, and reduces inflammation in atopic dry skin. International Journal of Dermatology, 2005. 44(2): p. 151-157.
85. Song, Y.H., et al., Muscle-specific expression of IGF-1 blocks angiotensin II-induced skeletal muscle wasting. Journal of Clinical Investigation, 2005. 115(2): p. 451-458.
86. Brink, M., et al., Angiotensin II stimulates gene expression of cardiac insulin-like growth factor I and its receptor through effects on blood pressure and food intake. Hypertension, 1999. 34(5): p. 1053-1059.
87. Ma, Y.W., et al., Angiotensin II stimulates transcription of insulin-like growth factor I receptor in vascular smooth muscle cells: Role of nuclear factor-kappa B. Endocrinology, 2006. 147(3): p. 1256-1263.
88. Kajstura, J., et al., IGF-1 overexpression inhibits the development of diabetic cardiomyopathy and angiotensin II-mediated oxidative stress. Diabetes, 2001. 50(6): p. 1414-1424.
89. Maggio, M., et al., Magnesium and anabolic hormones in older men. International Journal of Andrology, 2011. 34(6): p. E594-E600.
90. Debold, A.J., Atrial Natriuretic Factor - a Hormone Produced by the Heart. Science, 1985. 230(4727): p. 767-770.
91. Sultanian, R., Y.M. Deng, and S. Kaufman, Atrial natriuretic factor increases splenic microvascular pressure and fluid extravasation in the rat. Journal of Physiology-London, 2001. 533(1): p. 273-280.
92. Abell, T.J., et al., Atrial Natriuretic Factor Inhibits Proliferation of Vascular Smooth-Muscle Cells Stimulated by Platelet-Derived Growth-Factor. Biochemical and Biophysical Research Communications, 1989. 160(3): p. 1392-1396.
93. Bianciotti, L.G., et al., Atrial-Natriuretic-Factor Enhances Induced Salivary Secretion in the Rat. Regulatory Peptides, 1994. 49(3): p. 195-202.
94. Caisey, L., et al., Influence of age and hormone replacement therapy on the functional properties of the lips. Skin Research and Technology, 2008. 14(2): p. 220-225.
95. Mitteroecker, P., et al., The Morphometrics of "Masculinity" in Human Faces. Plos One, 2015. 10(2).
96. Jankowski, M., et al., Estrogen receptors activate atrial natriuretic peptide in the rat heart. Proceedings of the National Academy of Sciences, 2001. 98(20): p. 11765-11770.
97. Henesy, M.B., et al., Calcineurin regulates homologous desensitization of natriuretic peptide receptor-A and inhibits ANP-induced testosterone production in MA-10 cells. PloS one, 2012. 7(8): p. e41711.
98. Palmer, B.F. and D.J. Clegg, An Emerging Role of Natriuretic Peptides: Igniting the Fat Furnace to Fuel and Warm the Heart. Mayo Clinic Proceedings, 2015. 90(12): p. 1666-1678.
99. Delcayre, C., et al., Cardiac aldosterone production and ventricular remodeling. Kidney International, 2000. 57(4): p. 1346-1351.
100. McGraw, A.P., et al., Aldosterone Increases Early Atherosclerosis and Promotes Plaque Inflammation Through a Placental Growth Factor-Dependent Mechanism. Journal of the American Heart Association, 2013. 2(1).
101. Ishizawa, K., et al., Aldosterone stimulates vascular smooth muscle cell proliferation via big mitogen-activated protein kinase 1 activation. Hypertension, 2005. 46(4): p. 1046-1052.
102. NIH-library. Spironolactone. Available from: [https:// https://]www.nlm.nih.gov/medlineplus/druginfo/meds/a682627.html.
103. Kamei, H. and K. Arakawa, A Case of Pseudo-Aldosteronism Due to Addiction of Jintan, a Mouth Refresher Popular among Japanese. Japanese Heart Journal, 1982. 23(4): p. 651-659.
104. Caroccia, B., et al., GPER-1 and estrogen receptor-β ligands modulate aldosterone synthesis. Endocrinology, 2014. 155(11): p. 4296-4304.
105. Goncharov, N., et al., Effects of short-term testosterone administration on variables of the metabolic syndrome, in particular aldosterone. Hormone molecular biology and clinical investigation, 2012. 12(2): p. 401-406.
106. Klebanoff, C.A., et al., Retinoic acid controls the homeostasis of pre-cDC-derived splenic and intestinal dendritic cells. Journal of Experimental Medicine, 2013. 210(10): p. 1961-1976.
107. Chen, T.H.P., et al., Epicardial induction of fetal cardiomyocyte proliferation via a retinoic acid-inducible trophic factor. Developmental Biology, 2002. 250(1): p. 198-207.
108. Halevy, O. and O. Lerman, Retinoic Acid Induces Adult Muscle-Cell Differentiation Mediated by the Retinoic Acid Receptor-Alpha. Journal of Cellular Physiology, 1993. 154(3): p. 566-572.
109. Kautsky, M.B., P. Fleckman, and B.A. Dale, Retinoic acid regulates oral epithelial differentiation by two mechanisms. J Invest Dermatol, 1995. 104(4): p. 546-53.
110. Hong, W.K., et al., 13-Cis-Retinoic Acid in the Treatment of Oral Leukoplakia. New England Journal of Medicine, 1986. 315(24): p. 1501-1505.
111. Abbott, B.D., M.W. Harris, and L.S. Birnbaum, Etiology of Retinoic Acid-Induced Cleft-Palate Varies with the Embryonic Stage. Teratology, 1989. 40(6): p. 533-553.
112. Marzolla, V., et al., The role of the mineralocorticoid receptor in adipocyte biology and fat metabolism. Molecular and Cellular Endocrinology, 2012. 350(2): p. 281-288.
113. Murholm, M., et al., Retinoic acid has different effects on UCP1 expression in mouse and human adipocytes. Bmc Cell Biology, 2013. 14.
114. Xia, Q., et al., Effects of ghrelin on the proliferation and secretion of splenic T lymphocytes in mice. Regulatory Peptides, 2004. 122(3): p. 173-178.
115. Leite-Moreira, A.F., A. Rocha-Sousa, and T. Henriques-Coelho, Cardiac, skeletal, and smooth muscle regulation by ghrelin. Vitamins and Hormones, Ghrelin, 2008. 77: p. 207-+.
116. Ohta, K., et al., Expression and Possible Immune-regulatory Function of Ghrelin in Oral Epithelium. Journal of Dental Research, 2011. 90(11): p. 1286-1292.
117. Kellokoski, E., et al., Estrogen replacement therapy increases plasma ghrelin levels. The Journal of Clinical Endocrinology & Metabolism, 2005. 90(5): p. 2954-2963.
118. Greenman, Y., et al., Testosterone is a strong correlate of ghrelin levels in men and postmenopausal women. Neuroendocrinology, 2008. 89(1): p. 79-85.
119. Ueno, H., et al., Ghrelin: a gastric peptide that regulates food intake and energy homeostasis. Regulatory Peptides, 2005. 126(1-2): p. 11-19.
120. Klabunde, R.E., Cardiovascular physiology concepts. 2nd ed. 2012, Philadelphia: Wolters Kluwer Health/Lippincott Williams & Wilkins. xi, 243 p.
121. Kumar, P., et al., All-Trans Retinoic Acid and Sodium Butyrate Enhance Natriuretic Peptide Receptor A Gene Transcription: Role of Histone Modification. Molecular Pharmacology, 2014. 85(6): p. 946-957.
122. Torsello, A., et al., Ghrelin plays a minor role in the physiological control of cardiac function in the rat. Endocrinology, 2003. 144(5): p. 1787-1792.
123. Milosevic, V.L., et al., Central effects of ghrelin on the adrenal cortex: a morphological and hormonal study. General Physiology and Biophysics, 2010. 29(2): p. 194-202.
124. Pereira-Terra, P., et al., Neuroendocrine factors regulate retinoic acid receptors in normal and hypoplastic lung development. Journal of Physiology-London, 2015. 593(15): p. 3301-3311.
125. Suzuki, M., et al., Spleen-Derived Growth-Factor, Sdgf-3, Is Identified as Keratinocyte Growth-Factor (Kgf). Febs Letters, 1993. 328(1-2): p. 17-20.
126. Tichelaar, J.W., W. Lu, and J.A. Whitsett, Conditional expression of fibroblast growth factor-7 in the developing and mature lung. J Biol Chem, 2000. 275(16): p. 11858-64.
127. Dou, C.Q., et al., Strengthening the Skin with Topical Delivery of Keratinocyte Growth Factor-1 Using a Novel DNA Plasmid. Molecular Therapy, 2014. 22(4): p. 752-761.
128. Rosenquist, T.A. and G.R. Martin, Fibroblast growth factor signalling in the hair growth cycle: Expression of the fibroblast growth factor receptor and ligand genes in the murine hair follicle. Developmental Dynamics, 1996. 205(4): p. 379-386.
129. Ishibashi, T., et al., Keratinocyte growth factor and its receptor messenger RNA expression in nasal mucosa and nasal polyps. Annals of Otology Rhinology and Laryngology, 1998. 107(10): p. 885-890.
130. Voelkel, N.F., R.W. Vandivier, and R.M. Tuder, Vascular endothelial growth factor in the lung. American Journal of Physiology-Lung Cellular and Molecular Physiology, 2006. 290(2): p. L209-L221.
131. Owen, J.L., et al., Up-regulation of matrix metalloproteinase-9 in T lymphocytes of mammary tumor bearers: role of vascular endothelial growth factor. J Immunol, 2003. 171(8): p. 4340-51.
132. Yano, K., L.F. Brown, and M. Detmar, Control of hair growth and follicle size by VEGF-mediated angiogenesis. Journal of Clinical Investigation, 2001. 107(4): p. 409-417.
133. Detmar, M., The role of VEGF and thrombospondins in skin angiogenesis. Journal of Dermatological Science, 2000. 24: p. S78-S84.
134. Lee, H.S., A. Myers, and J. Kim, Vascular endothelial growth factor drives autocrine epithelial cell proliferation and survival in chronic rhinosinusitis with nasal polyposis. Am J Respir Crit Care Med, 2009. 180(11): p. 1056-67.
135. Gkini, M.A., et al., Study of platelet-rich plasma injections in the treatment of androgenetic alopecia through an one-year period. J Cutan Aesthet Surg, 2014. 7(4): p. 213-9.
136. Bagchi, M., et al., Vascular endothelial growth factor is important for brown adipose tissue development and maintenance. Faseb Journal, 2013. 27(8): p. 3257-3271.
137. Hemila, H. and P. Louhiala, Vitamin C may affect lung infections. Journal of the Royal Society of Medicine, 2007. 100(11): p. 495-498.
138. Fisher, B.J., et al., Ascorbic acid attenuates lipopolysaccharide-induced acute lung injury. Critical Care Medicine, 2011. 39(6): p. 1454-1460.
139. Tsao, C.S., P.Y. Leung, and M. Young, Effect of Dietary Ascorbic-Acid Intake on Tissue Vitamin-C in Mice. Journal of Nutrition, 1987. 117(2): p. 291-297.
140. Willis, R.J. and C.C. Kratzing, Extracellular Ascorbic-Acid in Lung. Biochimica Et Biophysica Acta, 1976. 444(1): p. 108-117.
141. Kwack, M.H., et al., L-ascorbic acid 2-phosphate represses the dihydrotestosterone-induced dickkopf-1 expression in human balding dermal papilla cells. Exp Dermatol, 2010. 19(12): p. 1110-2.
142. Farris, P.K., Topical vitamin C: A useful agent for treating photoaging and other dermatologic conditions. Dermatologic Surgery, 2005. 31(7): p. 814-818.
143. Thornhill, S.M. and A.M. Kelly, Natural treatment of perennial allergic rhinitis. Altern Med Rev, 2000. 5(5): p. 448-54.
144. Ryan, A.S. and L.A. Goldsmith, Nutrition and the skin. Clinics in dermatology, 1996. 14(4): p. 389-406.
145. Shimoda, L.A. and G.L. Semenza, HIF and the Lung Role of Hypoxia-inducible Factors in Pulmonary Development and Disease. American Journal of Respiratory and Critical Care Medicine, 2011. 183(2): p. 152-156.
146. Imamura, Y., et al., HIF-2 alpha/ARNT complex regulates hair development via induction of p21(Waf1/Cip1) and p27(Kip1). Faseb Journal, 2014. 28(6): p. 2517-2524.
147. Nys, K., et al., Uncovering the role of hypoxia inducible factor-1 alpha in skin carcinogenesis. Biochimica Et Biophysica Acta-Reviews on Cancer, 2011. 1816(1): p. 1-12.
148. Shin, H.W., et al., Hypoxia-inducible Factor 1 Mediates Nasal Polypogenesis by Inducing Epithelial-to-Mesenchymal Transition. American Journal of Respiratory and Critical Care Medicine, 2012. 185(9): p. 944-954.
149. Forsythe, J.A., et al., Activation of vascular endothelial growth factor gene transcription by hypoxia-inducible factor 1. Molecular and Cellular Biology, 1996. 16(9): p. 4604-4613.
150. Tsuji, K., S. Kitamura, and H. Makino, Hypoxia-inducible factor 1 alpha regulates branching morphogenesis during kidney development. Biochemical and Biophysical Research Communications, 2014. 447(1): p. 108-114.
151. Rodriguez, J.A., et al., Vitamins C and E prevent endothelial VEGF and VEGFR-2 overexpression induced by porcine hypercholesterolemic LDL. Cardiovascular Research, 2005. 65(3): p. 665-673.
152. Muellner, M.K., et al., Vitamin C inhibits NO-induced stabilization of HIF-1 alpha in HUVECs. Free Radical Research, 2010. 44(7): p. 783-791.
153. Geesin, J.C., et al., Regulation of Collagen-Synthesis in Human Dermal Fibroblasts in Contracted Collagen Gels by Ascorbic-Acid, Growth-Factors, and Inhibitors of Lipid-Peroxidation. Experimental Cell Research, 1993. 206(2): p. 283-290.
154. Gomez-Roman, J.J., et al., Hormone expression and opioid receptors in fetal and adult lung. Archivos De Bronconeumologia, 2002. 38(8): p. 362-366.
155. Hastings, R.H., et al., Alveolar Epithelial-Cells Express and Secrete Parathyroid Hormone-Related Protein. American Journal of Respiratory Cell and Molecular Biology, 1994. 11(6): p. 701-706.
156. Hsu, Y.J., et al., Calcitonin-stimulated renal Ca2+ reabsorption occurs independently of TRPV5. Nephrology Dialysis Transplantation, 2010. 25(5): p. 1428-1435.
157. Wallach, S., et al., Effects of Calcitonin on Bone Quality and Osteoblastic Function. Calcified Tissue International, 1993. 52(5): p. 335-339.
158. Lando, M., L.A. Hoover, and G. Finerman, Stabilization of hearing loss in Paget's disease with calcitonin and etidronate. Arch Otolaryngol Head Neck Surg, 1988. 114(8): p. 891-4.
159. Samuelov, L., et al., Neural controls of human hair growth: calcitonin gene-related peptide (CGRP) induces catagen. Journal of dermatological science, 2012. 67(2): p. 153-155.
160. Esbrit, P. and J. Egido, The emerging role of parathyroid hormone-related protein as a renal regulating factor. Nephrology Dialysis Transplantation, 2000. 15(8): p. 1109-1111.
161. Martin, T.J., Osteoblast-derived PTHrP is a physiological regulator of bone formation. Journal of Clinical Investigation, 2005. 115(9): p. 2322-2324.
162. Karperien, M., et al., Parathyroid hormone related peptide mRNA expression during murine postimplantation development: Evidence for involvement in multiple differentiation processes. International Journal of Developmental Biology, 1996. 40(3): p. 599-608.
163. Skrok, A., et al., The Effect of Parathyroid Hormones on Hair Follicle Physiology: Implications for Treatment of Chemotherapy-Induced Alopecia. Skin pharmacology and physiology, 2015. 28(4): p. 213-225.
164. Fargeas, M.J., J. Fioramonti, and L. Bueno, Central Actions of Calcitonin on Body-Temperature and Intestinal Motility in Rats - Evidence for Different Mediations. Regulatory Peptides, 1985. 11(2): p. 95-103.
165. Kir, S., et al., Tumour-derived PTH-related protein triggers adipose tissue browning and cancer cachexia. Nature, 2014. 513(7516): p. 100-+.
166. Pulkkinen, K., S. Murugan, and S. Vainio, Wnt signaling in kidney development and disease. Organogenesis, 2008. 4(2): p. 55-9.
167. Dean, C.H., et al., Canonical Wnt signaling negatively regulates branching morphogenesis of the lung and lacrimal gland. Developmental Biology, 2005. 286(1): p. 270-286.
168. Maeda, K., N. Takahashi, and Y. Kobayashi, Roles of Wnt signals in bone resorption during physiological and pathological states. Journal of Molecular Medicine-Jmm, 2013. 91(1): p. 15-23.
169. Riccomagno, M.M., S. Takada, and D.J. Epstein, Wnt-dependent regulation of inner ear morphogenesis is balanced by the opposing and supporting roles of Shh. Genes & Development, 2005. 19(13): p. 1612-1623.
170. Rabbani, P., et al., Coordinated Activation of Wnt in Epithelial and Melanocyte Stem Cells Initiates Pigmented Hair Regeneration. Cell, 2011. 145(6): p. 941-955.
171. Kang, S., et al., Effects of wnt signaling on brown adipocyte differentiation and metabolism mediated by PGC-1 alpha. Molecular and Cellular Biology, 2005. 25(4): p. 1272-1282.
172. Mount, P.F. and D.A. Power, Nitric oxide in the kidney: functions and regulation of synthesis. Acta Physiol (Oxf), 2006. 187(4): p. 433-46.
173. Danziger, R.S., B.S. Zuckerbraun, and J.M. Pensler, Role of nitric oxide in the regulation of osteoblast metabolism. Plastic and Reconstructive Surgery, 1997. 100(3): p. 670-673.
174. Takumida, M. and M. Anniko, Functional significance of nitric oxide in the inner ear. In Vivo, 2004. 18(3): p. 345-350.
175. Wolf, R., et al., Nitric oxide in the human hair follicle: constitutive and dihydrotestosterone-induced nitric oxide synthase expression and NO production in dermal papilla cells. Journal of molecular medicine, 2003. 81(2): p. 110-117.
176. Gooi, J.H., et al., Calcitonin impairs the anabolic effect of PTH in young rats and stimulates expression of sclerostin by osteocytes. Bone, 2010. 46(6): p. 1486-1497.
177. Lopez-Herradon, A., et al., Inhibition of the canonical Wnt pathway by high glucose can be reversed by parathyroid hormone-related protein in osteoblastic cells. Journal of Cellular Biochemistry, 2013. 114(8): p. 1908-1916.
178. Tas, N., et al., The effect of calcitonin treatment on plasma nitric oxide levels in post-menopausal osteoporotic patients. Cell Biochemistry and Function, 2002. 20(2): p. 103-105.
179. Kalinowski, L., L.W. Dobrucki, and T. Malinski, Nitric oxide as a second messenger in parathyroid hormone-related protein signaling. Journal of Endocrinology, 2001. 170(2): p. 433-440.
180. Munoz, F.J., et al., Wnt-5 alpha increases NO and modulates NMDA receptor in rat hippocampal neurons. Biochemical and Biophysical Research Communications, 2014. 444(2): p. 189-194.
181. Yavropoulou, M.P. and J.G. Yovos, The role of the Wnt signaling pathway in osteoblast commitment and differentiation. Hormones (Athens), 2007. 6(4): p. 279-94.
182. Glass, D.A., et al., Canonical Wnt signaling in differentiated osteoblasts controls osteoclast differentiation. Developmental Cell, 2005. 8(5): p. 751-764.
183. Gul, M., et al., Effect of erythropoietin on liver regeneration in an experimental model of partial hepatectomy. International Journal of Surgery, 2013. 11(1): p. 59-63.
184. Zanjani, E.D., et al., Studies on the liver to kidney switch of erythropoietin production. J Clin Invest, 1981. 67(4): p. 1183-8.
185. Uslu, M., et al., Erythropoietin stimulates patellar tendon healing in rats. Knee, 2015. 22(6): p. 461-468.
186. Tong, Z.Z., et al., Promoter polymorphism of the erythropoietin gene in severe diabetic eye and kidney complications. Proceedings of the National Academy of Sciences of the United States of America, 2008. 105(19): p. 6998-7003.
187. Kapsoritakis, A.N., et al., Finger clubbing and erythropoietin serum levels in active IBD. Inflammatory bowel diseases, 2006. 12(6): p. 535-536.
188. Gursu, M., et al., Skin disorders in peritoneal dialysis patients: An underdiagnosed subject. World Journal of Nephrology, 2016. 5(4): p. 372.
189. Wang, L., et al., PPAR alpha and Sirt1 Mediate Erythropoietin Action in Increasing Metabolic Activity and Browning of White Adipocytes to Protect Against Obesity and Metabolic Disorders. Diabetes, 2013. 62(12): p. 4122-4131.
190. Ishii, T., et al., Hepatocyte Growth-Factor Stimulates Liver-Regeneration and Elevates Blood Protein Level in Normal and Partially Hepatectomized Rats. Journal of Biochemistry, 1995. 117(5): p. 1105-1112.
191. Zhang, J.Y., et al., HGF Mediates the Anti-inflammatory Effects of PRP on Injured Tendons. Plos One, 2013. 8(6).
192. Nakase, J., et al., Facilitated Tendon-Bone Healing by Local Delivery of Recombinant Hepatocyte Growth Factor in Rabbits. Arthroscopy-the Journal of Arthroscopic and Related Surgery, 2010. 26(1): p. 84-90.
193. Grierson, I., et al., Hepatocyte growth factor/scatter factor in the eye. Progress in Retinal and Eye Research, 2000. 19(6): p. 779-802.
194. Karnath, B., Digital clubbing: a sign of underlying disease. Hospital Physician, 2003. 26: p. 25-27.
195. Bell, L.N., et al., A central role for hepatocyte growth factor in adipose tissue angiogenesis. American Journal of Physiology-Endocrinology and Metabolism, 2008. 294(2): p. E336-E344.
196. Ayers, C.R., Plasma Renin Activity and Renin-Substrate Concentration in Patients with Liver Disease. Circulation Research, 1967. 20(6): p. 594-&.
197. Monnouchi, S., et al., The Roles of Angiotensin II in Stretched Periodontal Ligament Cells. Journal of Dental Research, 2011. 90(2): p. 181-185.
198. VanHaeringen, N.J., The renin-angiotensin system in the human eye. British Journal of Ophthalmology, 1996. 80(2): p. 99-100.
199. Ena, P., et al., High prevalence of cardiovascular diseases and enhanced activity of the renin-angiotensin system in psoriatic patients. Acta cardiologica, 1984. 40(2): p. 199-205.
200. Koh, S.L., et al., Blockade of the renin-angiotensin system improves the early stages of liver regeneration and liver function. Journal of Surgical Research, 2013. 179(1): p. 66-71.
201. Suh, J.G., et al., Sequence analysis, tissue expression and chromosomal localization of a mouse secreted superoxide dismutase gene. Molecules and Cells, 1997. 7(2): p. 204-207.
202. Cui, Y.Y., et al., SOD Mimetic Improves the Function, Growth, and Survival of Small-Size Liver Grafts After Transplantation in Rats. Transplantation, 2012. 94(7): p. 687-694.
203. Okado-Matsumoto, A. and I. Fridovich, Subcellular distribution of superoxide dismutases (SOD) in rat liver: Cu,Zn-SOD in mitochondria. J Biol Chem, 2001. 276(42): p. 38388-93.
204. Kitamura, N., et al., Stress deprivation enhances manganese superoxide dismutase expression in the rat patellar tendon. International Journal of Molecular Medicine, 2004. 14(4): p. 537-543.
205. Behndig, A., et al., Superoxide dismutase isoenzymes in the human eye. Investigative Ophthalmology & Visual Science, 1998. 39(3): p. 471-475.
206. Li, C. and H.-M. Zhou, The role of manganese superoxide dismutase in inflammation defense. Enzyme research, 2011. 2011.
207. Petrovic, V., et al., Antioxidative defense and mitochondrial thermogenic response in brown adipose tissue. Genes and Nutrition, 2010. 5(3): p. 225-235.
208. MacLeod, A.K., et al., Expression and Localization of Rat Aldo-Keto Reductases and Induction of the 1B13 and 1D2 Isoforms by Phenolic Antioxidants. Drug Metabolism and Disposition, 2010. 38(2): p. 341-346.
209. Barski, O.A., S.M. Tipparaju, and A. Bhatnagar, The Aldo-Keto Reductase Superfamily and Its Role in Drug Metabolism and Detoxification. Drug Metabolism Reviews, 2008. 40(4): p. 553-624.
210. Ahmed, M.M.E., et al., Aldo-Keto Reductase-7A Protects Liver Cells and Tissues From Acetaminophen-Induced Oxidative Stress and Hepatotoxicity. Hepatology, 2011. 54(4): p. 1322-1332.
211. Artieda, M., et al., Tendon xanthomas in familial hypercholesterolemia are associated with a differential inflammatory response of macrophages to oxidized LDL. Febs Letters, 2005. 579(20): p. 4503-4512.
212. Huang, S.P., et al., Aldo-Keto Reductases in the Eye. Journal of Ophthalmology, 2010.
213. Shore, A.M., et al., Cold-Induced Changes in Gene Expression in Brown Adipose Tissue, White Adipose Tissue and Liver. Plos One, 2013. 8(7).
214. Wu, G.Y., et al., Glutathione metabolism and its implications for health. Journal of Nutrition, 2004. 134(3): p. 489-492.
215. Hung, L.K., et al., Local vitamin-C injection reduced tendon adhesion in a chicken model of flexor digitorum profundus tendon injury. J Bone Joint Surg Am, 2013. 95(7): p. e41.
216. Ganea, E. and J.J. Harding, Glutathione-related enzymes and the eye. Current Eye Research, 2006. 31(1): p. 1-11.
217. Niiyama, S. and H. Mukai, Reversible cutaneous hyperpigmentation and nails with white hair due to vitamin B-12 deficiency. European Journal of Dermatology, 2007. 17(6): p. 551-552.
218. Yang, H., et al., Effects of hepatocyte growth factor on glutathione synthesis, growth, and apoptosis is cell density-dependent. Experimental Cell Research, 2008. 314(2): p. 398-412.
219. Valdes-Arzate, A., et al., Hepatocyte growth factor protects hepatocytes against oxidative injury induced by ethanol metabolism. Free Radical Biology and Medicine, 2009. 47(4): p. 424-430.
220. Li, H., et al., HGF protects rat mesangial cells from high-glucose-mediated oxidative stress. American Journal of Nephrology, 2006. 26(5): p. 519-530.
221. Petrash, J.M., All in the family: aldose reductase and closely related aldo-keto reductases. Cellular and Molecular Life Sciences, 2004. 61(7-8): p. 737-749.
222. Hussein, A.M., et al., Effects of combined erythropoietin and epidermal growth factor on renal ischaemia/reperfusion injury: a randomized experimental controlled study. Bju International, 2011. 107(2): p. 323-328.
223. Zelko, I.N. and R.J. Folz, Extracellular superoxide dismutase functions as a major repressor of hypoxia-induced erythropoietin gene expression. Endocrinology, 2005. 146(1): p. 332-340.
224. d'Uscio, L.V., L.A. Smith, and Z.S. Katusic, Erythropoietin Increases Expression and Function of Vascular Copper- and Zinc-Containing Superoxide Dismutase. Hypertension, 2010. 55(4): p. 998-1004.
225. Weydert, C.J. and J.J. Cullen, Measurement of superoxide dismutase, catalase and glutathione peroxidase in cultured cells and tissue. Nature Protocols, 2010. 5(1): p. 51-66.
226. Chang, Q. and J.M. Petrash, Disruption of aldo-keto reductase genes leads to elevated markers of oxidative stress and inositol auxotrophy in Saccharomyces cerevisiae. Biochimica Et Biophysica Acta-Molecular Cell Research, 2008. 1783(2): p. 237-245.
227. Vlahakos, D.V., et al., Renin-Angiotensin System Stimulates Erythropoietin Secretion in Chronic-Hemodialysis Patients. Clinical Nephrology, 1995. 43(1): p. 53-59.
228. Yayama, K., et al., Angiotensin-converting enzyme inhibitor enhances liver regeneration following partial hepatectomy: Involvement of bradykinin B-2 and angiotensin AT(1) receptors. Biological & Pharmaceutical Bulletin, 2007. 30(3): p. 591-594.
229. Murakami, E., et al., Blood-Pressure Elevation Caused by Inhibition of Brain Glutathione-Reductase. Journal of Hypertension, Vol 7, Suppl 6, 1989: p. S24-S25.
230. Zhong, G., Zhong Yao Xue [Chinese Materia Medica]. . 2012, Beijing: China Press of Traditional Chinese Medicine
231. Hong, M., et al., The Decline of Atrial-Natriuretic-Peptide (Anp) Gene-Expression in Older Rats and the Effects of Ginsenoside on Anp Gene-Expression. Comparative Biochemistry and Physiology B-Biochemistry & Molecular Biology, 1992. 101(1-2): p. 35-39.
232. Kim, S.H., et al., Enhancement of 1,25-dihydroxyvitamin D3- and all-trans retinoic acid-induced HL-60 leukemia cell differentiation by Panax ginseng. Biosci Biotechnol Biochem, 2009. 73(5): p. 1048-53.
233. Xu, Z.W., et al., The effects of ginsenoside Rb1 on endothelial damage and ghrelin expression induced by hyperhomocysteine. Journal of Vascular Surgery, 2011. 53(1): p. 156-164.
234. Mao, N., et al., Ginsenoside Rg1 protects mouse podocytes from aldosterone-induced injury in vitro. Acta Pharmacologica Sinica, 2014. 35(4): p. 513-522.
235. Kimura, Y., et al., Effects of ginseng saponins isolated from Red Ginseng roots on burn wound healing in mice. British Journal of Pharmacology, 2006. 148(6): p. 860-870.
236. Zheng, K.Y., et al., Flavonoids from Radix Astragali induce the expression of erythropoietin in cultured cells: a signaling mediated via the accumulation of hypoxia-inducible factor-1alpha. J Agric Food Chem, 2011. 59(5): p. 1697-704.
237. Zhang, Y., et al., Radix Astragali extract promotes angiogenesis involving vascular endothelial growth factor receptor-related phosphatidylinositol 3-kinase/Akt-dependent pathway in human endothelial cells. Phytother Res, 2009. 23(9): p. 1205-13.
238. Ma, J., A. Peng, and S.Y. Lin, Mechanisms of the therapeutic effect of astragalus membranaceus on sodium and water retention in experimental heart failure. Chinese Medical Journal, 1998. 111(1): p. 17-23.
239. Chao, C.C., R.D. Brown, and L.J. Deftos, Seasonal Levels of Serum Parathyroid-Hormone, Calcitonin and Alkaline-Phosphatase in Relation to Antler Cycles in White-Tailed Deer. Acta Endocrinologica, 1984. 106(2): p. 234-240.
240. Barling, P.M., et al., Expression of PTHrP and the PTH/PTHrP receptor in growing red deer antler. Cell Biology International, 2004. 28(10): p. 661-673.
241. Lee, M.-R., et al., Hemopoietic effect of extracts from four parts of deer antler on phenylhydrazine-induced hemolytic anemia in female rats. Journal of the Korean Society of Food Science and Nutrition, 2009. 38(12): p. 1718-1723.
242. Chen, X., et al., [Protective effects of peptides from velvet antler of Cervus nippon on acute ischemic myocardial injury in rats]. Zhongguo Zhong Yao Za Zhi, 2009. 34(15): p. 1971-4.
243. Li, Z.H., W.H. Zhao, and Q.L. Zhou, [Experimental study of velvet antler polypeptides against oxidative damage of osteoarthritis cartilage cells]. Zhongguo Gu Shang, 2011. 24(3): p. 245-8.
244. Bensky, D., A. Gamble, and T.J. Kaptchuk, Chinese herbal medicine: materia medica. 1993: Eastland Press.
245. Shi P, Z.W., Li XM., Cynomorium Songaricum polysaccharide improves osteoporosis in ovariectomized rats. Journal of Third Military Medical University 2015. 37: p. 2360-2363
246. Yu, F.R., et al., Effects of a Flavonoid Extract from Cynomorium songaricum on the Swimming Endurance of Rats. American Journal of Chinese Medicine, 2010. 38(1): p. 65-73.
247. Hua, Z., et al., [Effect of angelica polysaccharides co-erythropoietin on JAK2/STAT5 signal transduction pathway in hematopoietic stem/progenitor cells]. Zhongguo Zhong Yao Za Zhi, 2009. 34(24): p. 3268-71.
248. Bi, C.W.C., et al., Fo Shou San, an Ancient Herbal Decoction Prepared from Rhizoma Chuanxiong and Radix Angelicae Sinensis, Stimulates the Production of Hemoglobin and Erythropoietin in Cultured Cells. Planta Medica, 2010. 76(14): p. 1525-1529.
249. Wang, Y., et al., Experimental studies on the HGF-inducing effect of L-cells treated with Angelica polysaccharide. Acta Anatomica Sinica, 1995. 27(1): p. 69-74.
250. Chiu, P.Y., et al., Dang-Gui Buxue Tang protects against oxidant injury by enhancing cellular glutathione in H9c2 cells: Role of glutathione synthesis and regeneration. Planta Medica, 2007. 73(2): p. 134-141.
251. Pu, X.Y., et al., Polysaccharides from Angelica and Astragalus exert hepatoprotective effects against carbon-tetrachloride-induced intoxication in mice. Canadian Journal of Physiology and Pharmacology, 2015. 93(1): p. 39-43.
252. Zhou, G.F., et al., Kinetic changes and antihypertensive effect of aqueous extract of Danggui (Angelica sinensis radix) after stir-fry processing. Food Science and Biotechnology, 2013. 22(3): p. 765-771.
253. Wen, Y.X., et al., Angelica Sinensis Polysaccharides Stimulated UDP-Sugar Synthase Genes through Promoting Gene Expression of IGF-1 and IGF1R in Chondrocytes: Promoting Anti-Osteoarthritic Activity. Plos One, 2014. 9(9).
254. Yu, H., et al., [Effect of Cordyceps sinensis on the expression of HIF-1alpha and NGAL in rats with renal ischemia-reperfusion injury]. Zhong Nan Da Xue Xue Bao Yi Xue Ban, 2012. 37(1): p. 57-66.
255. Singh, M., et al., Cordyceps sinensis Increases Hypoxia Tolerance by Inducing Heme Oxygenase-1 and Metallothionein via Nrf2 Activation in Human Lung Epithelial Cells. Biomed Research International, 2013.
256. Bhardwaj, A., et al., HPTLC Based Chemometrics of Medicinal Mushrooms. Journal of Liquid Chromatography & Related Technologies, 2015. 38(14): p. 1392-1406.
257. Yang, J.Y., et al., Effects of exopolysaccharide fraction (EPSF) from a cultivated Cordyceps sinensis fungus on c-Myc, c-Fos, and VEGF expression in B16 melanoma-bearing mice. Pathology Research and Practice, 2005. 201(11): p. 745-750.
258. Shi, S.G., et al., Overexpression of l-galactono-1, 4-lactone dehydrogenase (GLDH) in Lanzhou lily (Lilium davidii var. unicolor) via particle bombardment-mediated transformation. In Vitro Cellular & Developmental Biology-Plant, 2012. 48(1): p. 1-6.
259. Jiang, Y., et al., Wolfberry Water Soluble Phytochemicals Down-Regulate ER Stress Biomarkers and Modulate Multiple Signaling Pathways Leading To Inhibition of Proliferation and Induction of Apoptosis in Jurkat Cells. J Nutr Food Sci, 2011. S2.
260. Yi, R., X.M. Liu, and Q. Dong, A Study of Lycium Barbarum Polysaccharides (Lbp) Extraction Technology and Its Anti-Aging Effect. African Journal of Traditional Complementary and Alternative Medicines, 2013. 10(4): p. 171-174.
261. Li QY, L.Z., Jiang YN., Formulas of Traditional Chinese Medicine. . 1998, Beijing: Academic Press
262. Takeda, H., et al., Rikkunshito and Ghrelin Secretion. Current Pharmaceutical Design, 2012. 18(31): p. 4827-4838.
263. Shen, D.Z., et al., [Effects of Yiguanjian Decoction on liver cirrhosis formation:a differential proteomics study in rats]. Zhong Xi Yi Jie He Xue Bao, 2010. 8(2): p. 158-67.
264. Xia, B.J., et al., The effects of Liuwei Dihuang on canonical Wnt/beta-catenin signaling pathway in osteoporosis. Journal of Ethnopharmacology, 2014. 153(1): p. 133-141.
265. Ren, D., Lin Chuang Shi Yong Zhong Cheng Yao [Practical Clinical Chinese Medicine Formulas]. 2002, Beijing: People's Health Publishing House
266. Liu, M.J., et al., Effects of zuogui pill (see text) on Wnt singal transduction in rats with glucocorticoid-induced osteoporosis. J Tradit Chin Med, 2011. 31(2): p. 98-102.
267. Xiong, X.J., et al., Shenqi pill, a traditional Chinese herbal formula, for the treatment of hypertension: A systematic review. Complementary Therapies in Medicine, 2015. 23(3): p. 484-493.
268. Belefant-Miller, H. and S.C. Grace, Variations in Bran Carotenoid Levels within and between Rice Subgroups. Plant Foods for Human Nutrition, 2010. 65(4): p. 358-363.
269. Paine, J.A., et al., Improving the nutritional value of Golden Rice through increased pro-vitamin A content. Nature Biotechnology, 2005. 23(4): p. 482-487.
Document information

Published on 01/01/2017
Licence: CC BY-NC-SA license
